|
|
Trazando la Historia Genética del Hombre Moderno
Cavalli-Sforza, L. L., Menozzi, P., & Piazza, A.
The History and Geography of Human Genes (Princeton: Princeton
University Press, 1994)
![]()
Apunte sobre el conocimiento científico y la evolución humana. Carlos Rivera, profesor de Economía (12/02/2022).
La ciencia consigue
dar explicaciones cada vez más completas y precisas, basadas en evidencias
empíricas contrastables, y por tanto no utiliza la existencia de ningún ser
sobrenatural para explicar el origen del universo, del sistema
solar, de la tierra, de la vida o del hombre. El conocimiento está construyéndose
constantemente, lo que permite ir encajando la infinidad de piezas que componen
la realidad en este campo. Aunque hoy tenemos muchas certezas a las que se han
llegado por numerosas pruebas en distintos campos que se refuerzan, también hay
escasez de datos en algunos casos que obligan a plantearse distintas hipótesis
alternativas que tendrán que confirmarse con nuevos descubrimientos futuros.
El ser humano es un animal del orden de los primates, mamífero y vertebrado.
Su evolución ha sido fruto de la selección natural y ha estado condicionada por
factores muy diversos como las glaciaciones que modificaron el clima global, su
alimento, los depredadores y competidores (de distintas especies y de su misma
especie), las enfermedades, los parásitos, la selección sexual, la deriva génica,
etc. La expansión del hábitat de selva fragmentada y sabana favoreció inicialmente
su evolución hacia la locomoción bípeda, lo que permitió liberar las manos, mucho
antes del desarrollo del cerebro. Los caninos pudieron atrofiarse por no existir
presión selectiva hacia su desarrollo por tener otro tipo de defensas más eficaces
(utilización de ramas o palos y lanzamiento de objetos a distancia con precisión).
Por otro lado, la reducción de los caninos pudo implicar un menor nivel de agresión
física entre los miembros del grupo, favoreciendo su cohesión. Las muelas se desarrollaron
para adaptarse a una dieta que incorporaba semillas y frutos secos a una alimentación
principalmente vegetariana. El desarrollo progresivo de la inteligencia unido a la
habilidad para usar cada vez con más destreza las manos permitió la elaboración y
uso de herramientas inicialmente primitivas (posiblemente de madera, hueso y con
seguridad de piedra), que facilitó el aprovechamiento de recursos animales como
carroñero o cazador. El sistema de refrigeración consistente en multitud de glándulas
sudoríparas en una piel con poco pelo permitió a nuestros antepasados aprovechar
recursos en las horas centrales del día, ofreciendo poca superficie de exposición al
sol por la postura bípeda y asimismo protegidos por el pelo de la cabeza. El aumento
del tamaño corporal, de la fuerza física, del volumen cerebral unido a la reducción
del aparato digestivo y la acumulación de conocimiento que permitió elaborar
herramientas cada vez más sofisticadas dio a nuestros antepasados una ventaja cada vez
mayor sobre otros depredadores y sobre sus presas facilitando su expansión por
diferentes hábitats de distintos continentes evolucionando en distintas especies.
Posteriormente, la competencia entre grupos humanos, seleccionó a aquellos
grupos más cooperativos, complejos socialmente, innovadores, mejor comunicados, etc.
La colaboración entre grupos numerosos pudo ser favorecida por culturas, mitos o
creencias compartidas. El Homo sapiens moderno, que evolucionó en África, finalmente
se expandió por todo el planeta, contribuyendo probablemente a la extinción de algunas
de las especies humanas existentes (Homo neanderthalensis, homínido
de Denisova, Homo erectus, Homo
luzonensis y Homo floresiensis) e
hibridándose e incorporando, al menos en los dos primeros casos,
algunos de sus genes.
Resumen
En el siguiente trabajo se recopila abundante información que permite establecer un árbol de
relaciones genéticas del género humano. Conviene tener en cuenta que el vertiginoso avance de la
genética está matizando en muchos casos y contradiciendo en otros las conclusiones sacadas
en el pasado ya que hoy se dispone de gran cantidad de información y se cuenta con modelos
estadísticos y medios tecnológicos mucho más avanzados que cuando se realizó esta publicación.
En concreto hoy sabemos que la hibridación entre distintas especies humanas se ha producido
en múltiples ocasiones, así como los casos de mezcla entre humanos modernos. Esto implica
que el arbol filogenético de los humanos modernos en realidad es mucho más complejo y tiene
numerosas conexiones entre distintas ramas.
Esta masiva compilación de datos genéticos de las poblaciones del mundo, que documenta las
similitudes y diferencias genéticas, demuestra que las "razas"
existen a pesar de que simultáneamente se niega la utilidad del concepto.
Mientras se hace esto, el libro entrega mucha información de gran utilidad
sobre las similitudes y diferencias en las frecuencias genéticas entre las
poblaciones del mundo. Los datos son presentados en muchos y valiosos formatos
que incluyen gráficos, mapas, dendogramas (árboles
genealógicos) y diagramas de los principales componentes. La interpretación
generalmente presume neutralidad para los varios tipos de genes, y muchas
conclusiones interesantes son extraídas de la historia evolutiva de varias
poblaciones alrededor del mundo. Árbol
filogenético de los animales Árbol
filogenético de los cordados Árbol
filogenético de los mamíferos Árbol
filogenético de los primates Árbol
evolutivo de los homininos Árbol filogenético de
los humanos Gráfico de distancias genéticas Tabla de Contenidos Los gráficos de
las Distancias Genéticas 3 Árboles
genealógicos humanos 5 La separación
africana vs todas las otras es primordial 6 Otras
separaciones en el árbol 9 Otros interesantes
hallazgos en las distribuciones mundiales de genes 17 Conclusiones
sobre los Componentes Principales 17 Razas 22 Enfermedades y
Frecuencias Genéticas 24 Los capítulos
regionales 27 Asia 28 Europa 29 África 30 América 34 Conclusiones 35 El nuevo libro The History and Geography of
Human Genes de Cavalli-Sforza, Menozzi, & Piazza (1994) es una amplia colección de lo que se conoce
sobre la geografía y la historia de los genes humanos. Será un trabajo
fundamental en lo que a análisis raciales se refiere (a pesar que los autores
no lo describirían de ese modo). La mitad final del libro tiene un atlas que
muestra la distribución de un gran número de genes en cada continente, y en el
mundo entero. También hay un
amplio conjunto de gráficos que dan información sobre frecuencias de alelos
para muchos genes y poblaciones. Esta base de datos está compilada a partir del
examen de 2.900 artículos de 136 revistas (a pesar que solo 777 involucraban
datos no publicados, enumerados en las referencias). Es el cartografiado e
interpretación de esta inmensa cantidad de datos lo que hace el libro tan
impresionante y valioso. Desafortunadamente, la compilación de datos ya está de
algún modo obsoleta debido a que solo llega hasta 1986 (pág. 25). El libro está
organizado sobre una base geográfica, con un capítulo introductorio, un
capítulo sobre la distribución mundial de los genes y luego capítulos separados
sobre África, Asia, Europa, Oceanía (Australia, Nueva Guinea y las islas del
Pacífico) y América. El capítulo introductorio es una valiosa compilación de
material sobre genética, antropología, arqueología, evolución humana y los
métodos de genética cuantitativa. Los autores reconocen que el libro será usado
por gente que no son expertos en todos estos campos.
El material introductorio es útil para aquellos que carecen de capacitación en
uno o más de estos campos. Sin embargo, el especialista probablemente aprenderá
poco de las secciones de su propia especialidad. Una breve sección sobre el
"fracaso científico del concepto de las razas humanas" intenta
esquivar las críticas hacia los autores por atreverse a estudiar el problema de
cómo las frecuencias genéticas difieren alrededor del mundo (a pesar que
probablemente sí refleja las verdaderas creencias de los autores). Mucho del
libro parece contradecir las ideas anti-raza en esta sección, pero es un
efectivo argumento contra las más superficiales ideas que tienen las personas
sobre las razas. La última mitad del
primer capítulo contiene discusiones útiles sobre asuntos tales como el
problema de identificar poblaciones, y algunos de los métodos que serán usados
en el resto del libro. Mayor detalle, y un esfuerzo para colocar los métodos en
el más simple de los lenguajes, habrían sido útil para
el lector común y corriente. El segundo capítulo
resume los datos sobre la distribución mundial de los genes. El marco teórico
básico en este libro es que las frecuencias genéticas son determinadas por la
deriva. La descendencia hereda aleatoriamente los genes de sus padres. El niño
tiene un 50 % de oportunidades de heredar cualquier gen de cada progenitor.
Muchos quienes no están acostumbrados a pensar en términos genéticos piensan
que cualquier rasgo influenciado por los genes debe ser rígidamente heredado de
los padres. En la actualidad, debido a la herencia aleatoria de los genes, la
genética proporciona una teoría de la diversidad humana, y ayuda a explicar por
qué los vástagos generalmente son distintos (Rowe
1994). En poblaciones pequeñas y de algún modo aisladas, tales como los humanos
que se cree vivieron durante épocas prehistóricas, las frecuencias genéticas
cambian apreciablemente, pero de forma aleatoria, de generación en generación.
Sobre muchas generaciones (a lo mejor unos 100 mil años) las poblaciones
diferentes desarrollan frecuencias de genes diferentes. Los gráficos de Distancias Genéticas Como se mencionó,
una contribución del libro es el extenso gráfico de frecuencias genéticas. La
sección mundial está basada sobre datos de 120 alelos de 42 poblaciones (a
pesar que los datos no estaban disponibles para todos los alelos de todas las
poblaciones). Una cierta cantidad de promedio de datos de diferentes fuentes
fue necesaria para obtener las frecuencias genéticas relevantes, y los datos
son a menudo para grupos nacionales en Europa (como los ingleses o los
daneses), y grupos de tribus o regiones en otras partes del mundo.
Desafortunadamente, tras combinar varias poblaciones, hay más datos que
pudieron ser fácilmente incluidos. Son de gran ayuda los mapas al final del
libro que permiten al lector obtener una rápida visión de cómo todos los alelos
están distribuidos. Árboles genealógicos humanos Un método para
simplificar los datos es calcular las medidas de la distancia genética. Una de
las características más interesantes del libro son las numerosas matrices de
distancia genética que incluye, permitiendo ver cuán estrechamente están
relacionadas varias poblaciones. Ante varios ajustes estadísticos, estas son la
suma de los cuadrados de las diferencias en frecuencias promedio en los
numerosos genes. Bajo un modelo de deriva aleatoria, la distancia debiera ser a
grandes rasgos, proporcional al tiempo cuando las poblaciones se dividieron,
asumiendo que las poblaciones no diferían en tamaño, y que los genes no
estuvieron sujetos a una selección apreciable. Las distancias son generalmente
calculadas como distancia Fst, a pesar que las
distancias modificadas Nei son entregadas para una
muestra mundial de 42 poblaciones. La medida Fst
sería cero si todas las frecuencias genéticas disponibles fueran iguales, y 1
si ellas fueran completamente diferentes (por ejemplo, sí cada miembro de una
población siempre tuviera un alelo, cada miembro de la otra población carecería
de él). Estas medidas de
distancia genética son muy interesantes. En general uno no esperaría grandes
diferencias en frecuencias para un particular gen entre dos poblaciones, si
para la mayoría de los genes hubiera solo una pequeña diferencia en frecuencias
entre las dos poblaciones. Por ejemplo, cualquiera que arguyera que los
franceses y los alemanes difieren por razones genéticas, tendría que enfrentarse
con la evidencia presentada aquí que esas dos poblaciones son genéticamente muy
similares. El uso de la
distancia genética permite resumir los datos mundiales en una única matriz
triangular de 42 por 41. La mayor diferencia en el gráfico parece ser los 4.573
entre los pigmeos mbuti y los australianos (o sea,
los aborígenes australianos). En los gráficos los números son multiplicados por
10 mil para asunto de presentación. Por lo tanto, los Pigmeos parecen tener un
45.73 % de sus genes polimórficos diferentes de los australianos. De ese modo,
parece que para todos los pares de poblaciones humanas, la mayoría de los genes
serán comunes, incluso si uno escoge genes cuyos alelos difieren entre
individuos. Por supuesto, hay un número mucho mayor de genes que no parecen
tener variación conocida entre los humanos. La mayoría de los
datos son de genes que o son neutrales, o casi neutrales. Particularmente,
ninguno de los genes discutidos afectan
características genéticamente controladas como el color de piel, color de pelo,
color de ojos, forma o tamaño de la nariz. Todas estas características son
conocidas por diferir entre poblaciones. Del mismo modo, ninguno de los genes
estudiados es conocido por afectar directamente rasgos sociales tan importantes
como la inteligencia, la criminalidad, etc. (a pesar que muchos de esos rasgos
son conocidos por exhibir variabilidad genética, ver Bouchard,
Lykken, McGue, Segal, & Tellegen, 1990; Eaves, Eysenck, & Martin,
1989; Herrnstein & Murray, 1994; Miller, 1994a,b; Rowe 1994; Rushton,
1994). Los datos para los
ingleses, como población hacia la cual muchos norteamericanos trazan sus
orígenes, pueden ser usados para ilustrar la naturaleza de los datos. En la
muestra mundial de 42 poblaciones, la población más cercana a la Inglesa es la
Danesa (21), y la más distante los pigmeos mbuti de
Zaire (2.373). Las poblaciones africanas más importantes incluyen a los bantú
(2.288) y los africanos occidentales (1.487). Para propósitos de comparación,
las distancias genéticas entre los ingleses y los japoneses son de 1.244, entre
ellos y los chinos meridionales, 1.152, y entre estos y los amerindios del
Norte, 947. Estimaciones de errores estándar son proporcionados
por el método cruzado. Estas estimaciones indican que es imposible que el estudiar
genes neutrales adicionales cambie las conclusiones de que los ingleses tienen
frecuencias genéticas diferentes a aquellas poblaciones. Las distancias
genéticas entre los ingleses y otras poblaciones europeas son pequeñas. Las dos
mayores son 404 para los lapones y 340 para los sardos, dos poblaciones que
contribuyeron con pocos inmigrantes a los EE.UU. Con poblaciones europeas
mayores, 22 con los alemanes, las distancias son 24 con los franceses, 51 con
los italianos y hasta 204 con los Griegos. En comparación
con las distancias genéticas mucho mayores entre los bantú
y los africanos occidentales, o los japoneses, chinos meridionales y
amerindios, las poblaciones europeas efectivamente parecen similares unas con
otras. A menos que los genes que afectan los distintos tipos de comportamiento
tengan diferencias de frecuencias radicalmente diferentes a las de los genes
estudiados, las diferencias genéticas en comportamiento entre las poblaciones
europeas debieran ser pequeñas. Del mismo modo, las
diferentes poblaciones africanas occidentales son similares unas con otras. La
distancia promedio entre las varias tribus de África occidental es de 157, y
211 entre los grupos bantúes (pág. 184). Un bantú representativo para la
distancia africana occidental es de 188 (pág. 175). Dadas las enormes
distancias genéticas entre la mayoría de los europeos y la mayoría de los
africanos, y las similitudes dentro de las poblaciones que los esclavos e
inmigrantes Americanos extrajeron, parece razonable el dividir la mayoría de
los inmigrantes hacia Norteamérica de África o Europa en uno de los dos grupos
convencionales: negros y caucásicos. Es también lógico el pensar que grandes
diferencias genéticas aún existen entre las dos razas. Debido a que los dos
grupos originales difieren grandemente en color de piel, se espera que el color
de piel proporcione infomación sobre la probabilidad
de acarrear ciertos genes, incluso si las diferencias de población en
frecuencia son debido solo a la deriva (que es la
presunción principal para los genes discutida en este libro). Si los alelos han
sido sujetos a una selección relacionada con el clima, como ha sido argumentado
ser cierto para inteligencia y muchos aspectos del comportamiento (Lynn, 1991;
Miller 1991, 1994; Rushton, 1994), las diferencias
genéticas es posible que sean mayores. A pesar que a alguien le gustaría
argumentar que el conocimiento de las razas no es de utilidad al estimar la
probabilidad de que alguien tenga una característica en particular, la
aplicación del Teorema de Bayes demuestra lo
contrario. Las estadísticas Bayesianas demuestran que la estimación posterior
debiera ser un influyente promedio de la información sobre un individuo en
particular, y el promedio para la población de la cual él es miembro, con los
factores dependiendo de las relativas precisiones de la información sobre el
individuo y el grupo (Miller 1994c). Una matriz de
distancias genéticas contiene demasiados datos para ser entendidos rápidamente.
Los datos por lo tanto son presentados como dendogramas,
descritos como árboles en el libro. De este modo, en esta sección, y en el
resto del libro hay una extensa presentación de árboles. Las poblaciones que
están en las mismas ramas están estrechamente más relacionadas (como se muestra
en el gráfico de distancias genéticas). Los árboles son generalmente
interpretados como creados por la población humana original que se dividió y
subdividió. Una calibración a grandes rasgos es aplicada sobre las épocas
estimadas de movimiento de los modernos humanos fuera de África, y la
colonización de Australia y América. La longitud de las ramas representan el
relativo tiempo desde el cual que las poblaciones se separaron. Esto no siempre
es cierto, como los autores lo reconocen, debido a que las frecuencias
genéticas están más afectadas por la deriva en poblaciones pequeñas, y el flujo
de genes entre las poblaciones las hace más similares incluso aunque se hayan separado muchos años atrás. La separación africanos vs todos los demás es primordial Un argumento clave
de los resultados de los autores (pág. 78) muestra árboles de 42 poblaciones
usando frecuencias para 120 genes, con las distancias genéticas calculadas por
dos métodos diferentes. En los dos, la primera bifurcación separa a las
poblaciones africanas de las poblaciones no-africanas. Experimentos con métodos
cruzados (muestreando con reemplazos de la fuente de datos disponible, para
descubrir cuán sensitivas son las conclusiones frente a los cambios, en un
conjunto de genes examinados) demuestran que las poblaciones raíces africanas
(bantúes, nilo-saharianas, africanos occidentales y pigmeos mbuti)
se agrupan 83 a 84 veces de 100, mostrando una variación menor en los genes
estudiados que hacen poco probable el que cambien las conclusiones. Cuando las
42 poblaciones fueron agrupadas en 9 secciones, los africanos vs los
no-africanos fueron la primera división, y esto fue cierto para el 98 % de los
cruces (pág. 80). Se podría subrayar
que esto representa una modificación de los primeros resultados publicados
anteriormente (Edwards & Cavalli-Sforza, 1964, discutido por Cavalli-Sforza
y asociados, 1994, pág. 68), que ubicaba la primera bifurcación humana entre un
grupo caucasoide/negroide y todos los demás. Los
datos de frecuencia genética disponibles luego demostraron que los caucasoides
son más semejantes a los negroides que los mongoloides. El cambio por parte del
grupo de Cavalli-Sforza de agrupar a los mongoloides con los negroides (que
sería consistente si los humanos modernos que se originaron en Asia, seguidos
por una rama que se movía hacia el oeste, y luego subdividiéndose en grupos que
pasaron a África y Europa) se explica por el mucho mayor número de genes que
están disponibles ahora, más que por cualquier diferencia metodológica mayor. Incidentalmente, a
pesar que Cavalli-Sforza y sus asociados utilizan aquí la terminología de
africanos, europeos y asiáticos está claro que, a partir de las poblaciones
incluidas en cada grupo, lo que realmente ellos quieren decir es caucasoides,
negroides y mongoloides. Ellos se refieren a tomar tres poblaciones de cada
continente (pág. 68), pero los tres (pág. 68) muestran solo dos poblaciones del
continente de Europa (ingleses y lapones). Los turcos meridionales son
efectivamente localizados en Asia (o sea, son asiáticos), a pesar que son
caucasoides estrechamente semejantes a otras poblaciones europeas en
frecuencias genéticas. La exactitud y la claridad se verían mejoradas si los
términos raciales científicos estándar - caucasoides, negroides y mongoloides -
fueran utilizados, en vez de apropiarse de términos geográficos
tradicionalmente aplicados a los habitantes de continentes particulares,
europeos, africanos, asiáticos, y dándoles a estos términos un significado
nuevo e inusual. Particularmente, el libro usa más tarde "caucásico"
(Fig. 4.10.1 en la pág. 225) para referirse a alguien proveniente de las
montañas del Cáucaso, una utilización precisa, pero una que podría llevar a
confusión a un lector descuidado que solo mirara los árboles. Parece haber surgido
un acuerdo general sobre la primera ramificación en el árbol genealógico humano
la cual es entre los africanos y todos los demás. Esto ha sido demostrado por
numerosos métodos distintos. Como se mencionó anteriormente, es lo que se
esperaría si los humanos modernos se originaron en África, luego emigraron a
Oriente Próximo, y solo más tarde se dividieron en otras poblaciones. Nei & Roychoudhury
(1993) usando 26 poblaciones con los mismos genes para todas las poblaciones y
una metodología diferente (agrupación por vecindad) al grupo de Cavalli-Sforza,
halló que la primera bifurcación fue entre los africanos y los no-africanos, un
resultado que fue confirmado con 500 réplicas cruzadas. La ramificación fue
nuevamente africanos vs no-africanos, usando las mismas 26 poblaciones pero con
un método diferente de construcción de árbol (método de agrupación pareada sin
contrapeso más un promedio aritmético). En una prueba con 15 poblaciones pero
con más loci (33 loci y 131
alelos), la primera separación fue nuevamente entre africanos y no-africanos. Nei & Livshits
(1989) al examinar solo los tres grupos principales de africanos sub-saharianos
(mayoritariamente de Nigeria y Camerún), europeos (principalmente Gran Bretaña)
y asiáticos (principalmente japoneses) pudieron examinar 186 loci, que dieron suficientes datos para exámenes de
significancia estadística. Ellos descubrieron que la distancia entre los
africanos y los europeos era estadística y significativamente mayor que la
entre los europeos y los asiáticos, incluso a pesar que la geografía pone a
Gran Bretaña más cerca de Nigeria y Camerún, que de Japón. Mountain, Lin, Bowcock, &
Cavalli-Sforza (1993) enseñan un árbol, resultado de la utilización de 80
marcadores de ADN de 8 poblaciones. La primera brecha es entre africanos y
no-africanos. El árbol de Zhao & Lee (1989)
concuerda que la mayor diferencia genética es entre los africanos y todas las
otras poblaciones. Un estudio de
restricción del haplotipo de una enzima cercana al
gen de la b-globina demostró que "todas las poblaciones no-africanas
comparten un limitado número de haplotipos comunes,
cuando los africanos tienen predominantemente un haplotipo
diferente no encontrado en otras poblaciones. Un análisis de distancia genética
basado en estos polimorfismos nucleares de ADN indicaron que la mayor división
de las poblaciones humanas esta entre un grupo africano y un grupo
Euroasiático" (Wainscoat, Hill, Thein, Flint, Chapman, Weatherall,
Clegg & Higgs, 1989,
pág. 34). Torroni, Semino, Scozzari, Sirugo, Spedini, Abbas, Fellous, & santachiara Benerecetti (1990)
informaron sobre una aguda distinción entre africanos e italianos usando
marcadores sobre el cromosoma Y. Hammer (1994) ha
informado sobre un marcador del cromosoma Y (que implica heredabilidad
solo en línea masculina) que tenía una frecuencia de 0.74 en 611 africanos,
pero solo 0.7 en 192 europeos. Un árbol mostro que la primera separación fue
nuevamente entre africanos y no-africanos (a pesar que los Egipcios
fueron agrupados con los africanos). Conclusiones
similares han sido alcanzadas por otros investigadores utilizando otros
marcadores genéticos. Relethford & Harpending (1994) muestran que un árbol construido mediante
la utilización de la variación craniométrica, la
primera bifurcación fue entre los africanos y todos los demás. Otras bifurcaciones en el árbol Aunque la primera
separación en el árbol es clara y parece estar bien establecida, la segunda
separación es un poco sorprendente. Con el conjunto de mediciones de distancia
preferido (Fst), los no-africanos se separaron entre
australianos y todos los demás, y luego en asiáticos sudorientales y el resto.
Solo entonces los caucasoides se separan de los asiáticos nororientales,
asiáticos árticos y amerindios. Utilizando un método alternativo para calcular
las distancias genéticas - distancias Nei - los
no-africanos primero se separaron en un grupo Australiano y asiático
sudoriental, y un grupo caucasoide, asiático de otras
zonas y Americano. Luego los caucasoides se separaron de los asiáticos
nororientales, asiáticos Árticos y Americanos. Combinando las 42 poblaciones en
9 secciones (que incrementa el número de loci que
pueden ser utilizados y reduce la importancia de la deriva aleatoria), los
no-africanos son entonces separados en un grupo que combina a los australianos,
asiáticos del sudeste e isleños del Pacífico, y en un grupo que incluye a los
caucasoides, asiáticos del nordeste y Americanos. Los resultados aquí
son sorprendentes debido a que se descubre que genéticamente los asiáticos del
nordeste (incluyendo japoneses, coreanos y chinos septentrionales) y los
amerindios están relativamente más cerca de los geográficamente distantes
caucasoides, que de los asiáticos del sudeste, quienes están mucho más cerca.
Esto es algo que muchos podrían haber adivinado por la geografía o por la
semejanza física de las poblaciones. Interesantemente, una detallada inspección
de los árboles y de las matrices de distancia demuestran que los chinos
sudorientales (o sea, Hong Kong y alrededores) se agrupan con los asiáticos
sudorientales, como los filipinos, más que con los chinos septentrionales. Tal resultado no es
imposible. Uno podría imaginar que la antigua población de Oriente Próximo dio
luz a un grupo que emigró hacia el este, hasta el sudeste asiático, y luego a
Australia y Nueva Guinea. Más tarde la población de Oriente Proximo
podría haber dado a luz grupos que se transformaron en caucasoides, asiáticos
del nordeste y amerindios. Los autores
conducen experimentos cruzados (que en esencia repiten los cálculos con diferentes
conjuntos de genes, para ver cuán sensitivas las conclusiones son en relación
al particular conjunto de genes de los cuales tenemos datos). Las conclusiones
parecen cambiar dependiendo del conjunto de genes estudiados, y los autores
sugieren que uno no puede confiarse en el orden exacto de separación entre las
ramas que llevan a los caucasoides, asiáticos nororientales, asiáticos
sudorientales y las poblaciones australianas y de Nueva Guinea. Ellos atribuyen
mucho de esta poca certeza a un extenso flujo genético (mestizaje) entre el
nordeste y sudeste de Asia, que hace difícil el producir un árbol que encaje
bien los datos. La principal
alternativa a los extensos cálculos llevados a cabo por el grupo de
Cavalli-Sforza es otro conjunto de cálculos hechos por Nei
& Roychoudhury (1993). Como ya se mencionó, estos
cálculos concuerdan que la primera separación es entre los africanos y los
no-africanos. Sin embargo, ellos ubican la segunda separación entre los
caucasoides y los asiáticos principales (australianos, mongoloides y
Americanos). Los árboles que ellos construyen corresponden bastante a las razas
como han sido entendidas tradicionalmente, con sus poblaciones agrupadas en
árboles en grupos que son fácilmente reconocidos como negroides, caucasoides,
mongoloides, amerindios y australianos. La única diferencia de las razas
tradicionales es que la rama del árbol que lleva a los mongoloides también
incluye a los grupos australianos y de Nueva Guinea. Sin embargo, ellos están
en una rama separada. Nei & Roychoudhury
discuten por qué ellos obtienen un árbol distinto al de Cavalli-Sforza y
asociados, y concluyen que es porque ellos utilizan un método diferente para
construir árboles, agrupaciones por vecindad, mientras que Cavalli-Sforza y
asociados usan un método de vinculación promedio. Nei
& Roychoudhury presentan algunas razones
similares para preferir sus métodos alternativos. La mayoría de la
interpretación de los datos por Cavalli-Sforza y asociados está basada en la
deriva genética (o sea, los cambios aleatorios en las frecuencias genética que
suceden de una generación a otra). La idea implícita es que la mezcla de
poblaciones ha jugado un pequeño papel. Sin embargo, reconocen que la teoría
que observó diferencias en las frecuencias genéticas debido a la deriva es, al
menos en principio, comprobable. Por ejemplo, si no hay mezcla tras la
separación, todas las poblaciones que descienden de una misma población
original, debieran tener aproximadamente las mismas distancias genéticas de
varias poblaciones que descienden de otra población original (ver gráfico en
pág. 90). Esta condición no necesariamente se cumple si hay un apreciable flujo
genético entre poblaciones. En general, podríamos esperar que poblaciones
adyacentes intercambiaran genes, y que fueran más similares que las poblaciones
no-adyacentes. Ciertos métodos de
construcción producen árboles, en los que la longitud de sus ramas desde el
punto de origen indica cuánta separación genética ha ocurrido desde que las
poblaciones se separaron. Si las poblaciones están evolucionando a la misma
velocidad, todas las ramas de un punto en común debieran tener la misma
longitud. Muy a menudo esta condición no se cumple. Posiblemente la más
asombrosa excepción al patrón predicho es que la rama que lleva a los europeos
es a menudo relativamente corta. Uno de los más interesantes estudios
discutidos en el libro es uno que analizó solo un pequeño número de poblaciones
(incluyendo los chinos, europeos, dos poblaciones de pigmeos africanos y
Melanesios), pero coleccionaba datos de un gran número de alelos. Un árbol
construido usando estos datos enseñó una rama muy corta que nos lleva hacia los
europeos (pág. 91). Numerosas explicaciones fueron consideradas, pero la más
plausible fue la mezcla. Los cálculos demostraron que las frecuencias genéticas
europeas podrían ser explicadas bien por una mezcla de chinos con un menor
porcentaje de pigmeos. Obviamente, esto no es la verdadera historia racial de
los europeos (que son los más altos y de piel más clara de los grupos de la
muestra). Los pigmeos son ligeramente cercanos a otros africanos en la
frecuencia de sus genes medidos (el conjunto de frecuencias genéticas medidas
parece incluir ningún gen que afecte la estatura) de acuerdo a los datos en
este libro. La evidencia de que
las frecuencias genéticas europeas tienden a ser intermedias entre los
africanos y los chinos, es interesante para aquellos (incluyendo el autor de
este artículo) interesados en las diferencias de comportamiento entre las
razas. Rushton (1994) ha presentado evidencia que
sobre un amplio rango de características, incluyendo inteligencia y
comportamiento sexual, las razas se ordenan en mongoloides, caucasoides y
negroides. Él interpreta esto como evidencia para su teoría diferencial K,
mientras el autor de este artículo interpreta ese mismo patrón como evidencia
para su teoría de dedicación paterna (Miller, 1994a, b). Ambos han interpretado
el hecho de que tantas características tengan el mismo patrón como una
regularidad sistemática requiere una explicación. Esto fue explicado de forma
más fácil por un mecanismo evolutivo, que probablemente tomo la forma de un
factor relacionado al clima común, que produce diferencias debido a que los
africanos estén en el extremo tropical (o el extremo variable e impredecible en
la teoría de Rushton) y los mongoloides en el otro
extremo (el frío en el registro de Miller o lo predecible en el de Rushton) con los caucasoides al medio. Por supuesto, si las
frecuencias genéticas caucasoides son simplemente el resultado de la mezcla de
las otras dos familias, la regularidad debiera ser explicada en otra forma (a
pesar que las explicaciones de Miller y Rushton todavia podrían ser correctas). ¿Cómo las
frecuencias genéticas europeas llegan a ser intermedias entre los chinos y los
africanos? Parte de la explicación es simplemente geográfica. Los caucasoides
están localizados entre el territorio de los negroides y los mongoloides, y
presumiblemente han recibido genes de ambos grupos. El término caucasoide es utilizado en vez de meramente europeo debido
a que los caucasoides de Oriente Próximo y la India son quienes están mejor
localizados para intercambiar genes con negroides y mongoloides. Sin embargo, una
teoría de Ammerman & Cavalli-Sforza (1973), discutida en este libro (pág. 108)
proporciona un mecanismo para saber cómo los europeos pudieron llegar a ser
intermedios entre las frecuencias genéticas. Ellos concuerdan con la base de la
evidencia arqueológica y los gradientes de las frecuencias genéticas en Europa,
que la agricultura, tras emerger en Oriente Próximo, se difundió hacia Europa
por medio de la difusión démica. Por difusión démica se quiere decir que las antiguas poblaciones
agrícolas se expandieron gradualmente con cada nueva generación que emigraba
hacia Europa, con la velocidad promedio de un kilómetro por año. La alternativa
a este cuadro es que la técnica agrícola se difundió sin migraciones de
pueblos. Uno de los más
fascinantes trabajos consignados en el libro es la explicación de las
distribuciones de frecuencias genéticas, por medio de la hipótesis de la
difusión démica de la agricultura. Los autores
computan los primeros componentes principales para las frecuencias genéticas
europeas. Para aquellos no familiarizados con las estadísticas, el primer
componente principal es una única estadística que condensa la mayor información
posible sobre las frecuencias genéticas en un único número. Cuando se
cartografía, el componente se incrementa sistemáticamente con la distancia
desde Oriente Próximo. Esto se explica por el avance gradual de la población
agrícola de Oriente Próximo en Europa. Donde sea que hay un contacto entre dos
poblaciones, hay mestizaje, y los genes de las poblaciones europeas originales
gradualmente se difuminan en la población agrícola que avanza. Es una hipótesis
fascinante, y la utilización de los componentes principales para respaldarla es
ingenioso. Muchos genes individuales están
distribuidos como si hubieran sido importados por una población avanzando por
Europa desde Oriente Próximo, con la oleada gradualmente mezclándose más a
medida que la población intrusa se mestiza con los habitantes originales de
Europa. Tal cuadro
concuerda con lo que sabemos sobre los agricultores y pastores primitivos. Las
poblaciones nómades son típicamente de muy baja densidad, mientras que las
agrícolas pueden mantener densidades mucho más altas. Por lo tanto, un cambio
hacia la agricultura puede plausiblemente incrementar el índice de crecimiento
en la población, permitiendo que las densidades se incrementen rápidamente. Una
de las limitaciones al crecimiento demográfico en sociedades de pastores
nómades es la incapacidad de la madre para hacerse cargo de más de un vástago a
la vez. Esto evita que tenga el siguiente hijo (o que le permita sobrevivir)
hasta que el primero pueda caminar. Por lo tanto, los nacimientos están
espaciados en alrededor de cuatro años. En las poblaciones
agrícolas sedentarias, los nacimientos pueden ser más frecuentes, permitiendo
que la población crezca, al menos cuando hay tierra fértil adecuada para la
expansión. A medida que la población crece, las aldeas de vez en cuando crecen
tanto que se separan, con un grupo que las ha abandonado para establecer una
nueva aldea. Esta nueva aldea podría frecuentemente localizarse en una nueva
área sin colonizar. La agricultura
sedentaria es un modo de vida bastante diferente al nomadismo pastoral, y uno
que de cierta forma es físicamente más arduo. La evidencia de nómades
contemporáneos demuestra que ellos son reacios a adoptar la agricultura o un
medio de vida sedentario, mientras el pastoreo les proporcione el ingreso
adecuado. Pastores y agricultores sedentarios parecen haber vivido cerca unos
de otros, durante largos períodos de tiempo sin que los nómades adoptaran la
agricultura. También parece que mientras hubo cierto flujo genético entre tales
poblaciones, básicamente se mantuvieron separadas. De este modo, el
cuadro de Cavalli-Sforza y asociados, de difusión démica
de la agricultura es plausible. Ellos ilustran los cálculos demuestran que el
índice de avance observado (como ha sido medido en los sitios arqueológicos) es
cercano a lo que se esperaría de una expansión como la démica
(pág. 108). Además hay un
interés intrínseco ¿Cuál es la importancia de sí la agricultura en Europa fue
introducida por difusión démica o por difusión
cultural? Si fue efectivamente por un movimiento de pueblos, los actuales
habitantes de Europa son en gran medida, provenientes de Oriente Proximo, más que descendientes de los originales habitantes
Paleolíticos. Debido a que la agricultura asegura una más elevada densidad
demográfica, el impacto sobre las frecuencias genéticas sería bastante mayor a
causa de una invasión de agricultores como esa. Como se hizo notar
anteriormente, los datos sobre la frecuencia genética demuestran que las
frecuencias genéticas de los europeos parecen ser el resultado de una mezcla de
un tercio africano y dos tercios asiáticos. Mientras esta mixtura pudo ocurrir
por difusión directa en Europa, desde África o Asia (e indudablemente hubieron
flujos genéticos como esos), es más fácil entender que si los ancestros de los
europeos estuvieron originalmente en Oriente Próximo, posiblemente incluso
Israel (donde hay evidencia de una cultura sedentaria que almacenaba granos
silvestres, que podría fácilmente haber mutado a cultivar tales granos). Tal
población pudo haber estado recibiendo genes desde África, por medio del istmo
de Suez (y posiblemente a través del mar Rojo) y desde Asia. La evidencia de
difusión démica también arroja luz sobre el clima en
el cual los europeos evolucionaron. Es un lugar común en la psicología
evolutiva (también llamada sociobiología) que la
psicología humana (y el cuerpo) se le dio forma por medio de un extremadamente
largo período en el cual las personas eran pastores, y que probablemente fuimos
adaptados hacia el éxito reproductivo en lo que a menudo se denomina el medio
ambiente. Sin embargo, un pequeño vistazo demostrará que esos medios ambientes
variaron en diferentes partes del mundo y oscilaron desde lo tropical hasta el
frío de la Edad del Hielo en Eurasia. El autor de este
artículo ha argumentado en otros lugares (Miller 1994a, b) distintos a las
áreas tropicales, el alimento vegetal estaba disponible alrededor de una vez al
año. Se ha transformado en una generalización común que en las sociedades de
cazadores-recolectores, la mayoría de las calorías provengan de la recolección
y que la mayoría de la recolección fuera hecha por mujeres, y que el número
total de horas dedicadas a tal actividad era bajo (Lee, 1968). Sin embargo, un
examen de las sociedades estudiadas para establecer esta generalización
demuestra que eran sociedades típicamente tropicales. En tales áreas
tropicales, las mujeres podían recolectar suficiente comida para alimentarse a
ellas y sus hijos. La estrategia masculina óptima era dedicar todos sus
esfuerzos a la procreación con la mayor cantidad de mujeres posibles, y evitar
que otros hombres procrearan con las mujeres con las que él ha procreado. La
manutención de las mujeres y sus hijos no era tan necesaria para su éxito
reproductivo (debido a que ellas sobrevivirán de cualquier modo). En Eurasia, el
mayor problema era sobrevivir al del invierno cuando las frutas, arándanos,
insectos, huevos y animales hibernantes y migratorios
no estaban disponibles. Las soluciones comunes son el almacenamiento de
alimento (que lleva a la selección de la habilidad para obtener gratificación y
la inteligencia, ver Miller 1991), y la caza de grandes animales, tales como el
venado. Desafortunadamente, las mujeres no son efectivas cazadoras de grandes
animales (simplemente imaginen tratar de cazar mientras se acarrea a un bebé
llorando). De este modo, los hombres se transforman en los proveedores
principales de sus familias durante los inviernos. Las mujeres son entonces
seleccionadas para buscar y atraer hombres que las mantengan a ellas y a sus
hijos. Los hombres de Eurasia fueron seleccionados para formar fuertes lazos de
pareja y para tener rasgos de personalidad que llevaran a sostener y cuidar a
sus mujeres. Los ancestros de
los negroides eran africanos tropicales, y los ancestros de los mongoloides y
caucasoides provenían de las regiones de clima frío de Eurasia. Es más, para
explicar los mayores lazos de pareja de los mongoloides y otros atributos se
hace necesario argumentar que ellos evolucionaron en climas más fríos. Su
constitución compacta y otros rasgos son concuerdan con eso. Que los negroides
evolucionaron en el África tropical, y que los caucasoides y mongoloides
evolucionaron en la fría Eurasia es fácilmente aceptado. Sin embargo, algunos
han hallado difícil de creer que el medio ambiente en el que los caucasoides
evolucionaron fuera apreciablemente más tibio que el de los mongoloides,
especialmente debido a los registros de una Edad del Hielo en Europa.
Definitivamente era muy frío. Sus habitantes cazaban animales como el venado y
los mamuts lanudos. El modelo de
difusión démica hace probable que los ancestros de
los europeos modernos, no fueran mayoritariamente europeos de la Edad del
Hielo, sino habitantes Paleolíticos de Oriente Próximo. Las frecuencias
genéticas de los modernos europeos fueron formadas no solo por las frías
condiciones de la Europa de la Edad del Hielo, sino primariamente por las
condiciones en un Oriente Próximo de clima más tibio, posiblemente incluso en
Israel. Luego, las frecuencias genéticas aquí fueron influenciadas por los
genes que se diseminaron a través del istmo de Suez, desde África. ¿Qué sucedió con
los habitantes Paleolíticos originales de Europa? Fueron absorbidos dentro de
las poblaciones de agricultores que llegaron desde Oriente Próximo. Sin
embargo, la evidencia presentada en este libro sugiere que la población
existente que está más cercana a los europeos originales son los Vascos (pág. 276). Si la expansión de
los agricultores de Oriente Proximo afectó las
frecuencias genéticas en Europa, debió haber afectado las frecuencias genéticas
en las cuales esta agricultura pudo haberse difundido (págs. 221-222). Tiempo
después de que este libro fue publicado, Barbujani, Pilastro, Domenico & Renfrew
(1994), utilizaron datos de frecuencias genéticas para argüir que no solo las
frecuencias genéticas europeas sugieren una difusión démica
desde Oriente Proximo, sino que tal difusión démica también puede ser encontrada entre las áreas alguna
vez ocupadas por los hablantes de lenguas altaicas, y los asiáticos hablantes
de lenguas indoeuropeas y Elamo-Dravidianas, a pesar
que solo se halló débil evidencia entre los hablantes de lenguas
afro-asiáticas. Otros interesantes hallazgos sobre las distribuciones genéticas
mundiales Tras derivar los
árboles genealógicos, Cavalli-Sforza y asociados los compararon con la
distribución de las familias lingüísticas. Ellos concluyen que son similares.
Esto no es sorprendente debido a que lenguajes y genes, se asegura, se difunden
por repetidas divisiones entre las poblaciones, seguidas por la evolución
independiente de las frecuencias genéticas y los lenguajes. También, la gente
tiende a no casarse con aquellos que hablan lenguas diferentes, y las
diferencias lingüísticas se transforman en barreras para los flujos genéticos.
Debiera notarse que Nei & Roychoudhury
(1993), al construir sus árboles en un modo de una manera diferente,
encontraron una relación menos estrecha entre grupos genéticos y lenguajes. Conclusiones sobre los Componentes Principales Otra forma en que
la inmensa cantidad de datos en el gráfico de distancias genéticas puede ser
condensada, es calculando los componentes principales. En esencia, el primer
componente principal es un número que resuma tanta información sobre
frecuencias genéticas como sea posible. Después que esto sea hecho, un segundo
componente puede ser calculado para que resuma tanto como sea posible de la
información restante, y así sucesivamente. Los componentes principales no
siempre existen. Si la frecuencia de un alelo fuera completamente independiente
de las frecuencias de los otros alelos, los componentes principales no
existirían para ser calculados. Los componentes principales son utilizados en
otros campos. Por ejemplo, en psicología el primer componente principal de una
batería de exámenes es tradicionalmente llamado "g" (por habilidad
general), y es usualmente lo que el psicometrista
denomina inteligencia. Los dos primeros
componentes principales explican el 27 % y el 16 % de la variación
respectivamente (pág. 81). De este modo hay un alto grado de patrón en la
distribución de las frecuencias genéticas. Una técnica gráfica coloca las
poblaciones sobre un diagrama bidimensional con el primer componente principal
a lo largo de la base, y el segundo sobre el eje vertical. Como los autores lo
hacen notar, las poblaciones africanas están en el cuadrante derecho inferior,
y todos los caucasoides en el cuadrante derecho superior. Los tradicionalmente
llamados mongoloides están en el lado izquierdo del diagrama, junto a los
australianos y los de Nueva Guinea. Parece que cuando los datos modernos de
frecuencias genéticas son analizados con sofisticados y modernos métodos
estadísticos, producen algo que se parece mucho al tradicional concepto de
razas. La única excepción es que las poblaciones de Australia y Nueva Guinea
están en el medio del lado izquierdo, con poblaciones tradicionalmente
consideradas mongoloides sobre y debajo de ellas. Como en los árboles antes
mencionados (árboles y diagramas de componentes principales son meramente
diferentes modos de simplificar y presentar visualmente la misma información),
los mongoloides parecen caer en un grupo en la esquina superior izquierda
(incluyendo japoneses, coreanos, mongoles, ainus y grupos amerindios), y luego
en el cuadrante inferior izquierdo, otro grupo que incluye a los chinos
meridionales, Thai, indonesios, malayos, filipinos, etc.). Como un paréntesis
podemos decir, el lector podría notar que el segundo componente principal
parece dividir poblaciones, de algún modo mediante el área climática en la que
fueron encontrados, con los negroides, asiáticos meridionales, australianos y
habitantes de Nueva Guinea en el fondo del diagrama; y los caucasoides,
asiáticos del nordeste y grupos amerindios en la parte superior. Es posible que
los genes que jugaron un papel mayor en determinar el segundo componente, son los que están sujetos a la selección natural que de
algún modo se relaciona al clima (posiblemente a través de los efectos de las
enfermedades tropicales). El lector podría
darse cuenta que ciertas poblaciones que han contribuido fuertemente al
poblamiento de América, son cercanas unas con otras en el gráfico. En la
esquina superior derecha, los italianos, daneses, ingleses, Griegos e Iraníes,
son muy cercanos unos con otros. La referencia al capítulo sobre Europa muestra
que la mayoría de las otras poblaciones europeas (como los alemanes, franceses
y holandeses) que ayudaron a poblar América son muy similares a las poblaciones
cartografiadas aquí. En la esquina
inferior izquierda, las poblaciones bantú y africana
occidental resultan ser muy similares. El grupo europeo y el grupo africano
están casi tan alejados cómo es posible en el segundo componente. Mientras las
frecuencias genéticas estudiadas no afectan la apariencia de los individuos, es
plausible que si lo hicieran, la diferencia entre los europeos y los africanos
sería inmediatamente aparente, y que las palabras emergerían para describirlas.
Por supuesto, estas dos poblaciones están agudamente separadas por el color de
piel y otros aspectos de apariencia (debido a otros genes), y no es
sorprendente encontrar que esta diferencia en apariencia haya sido detectada.
Pueblos de los dos continentes originales son descritos como negros y blancos,
afroamericanos y euroamericanos, negros y caucásicos, etc. El segundo
componente principal hace un buen trabajo separando a los dos grupos de
poblaciones. Hay poblaciones que
yacen entre las dos secciones anteriormente descritas. Los Bereberes están casi
a medio camino en el segundo componente principal, y los san y africanos del
este mucho más cerca de otros africanos. Es muy posible que estos grupos
localizados en África reflejen diferentes grados de mezcla caucasoide
con la familia africana. Esta posibilidad con respecto a los
san es discutida en el capítulo africano. Debido a que estos grupos
contribuyeron relativamente poco al poblamiento de América, una impresión de
aguda distinción entre aquellos de ancestro no mezclado que actualmente existe
en el Viejo Mundo podría ser creada. Uno podría
preguntar qué pueden indicar los mapas de componentes principales. Los autores
argumentan que cuando dos poblaciones se mezclan, todas las frecuencias
genéticas cambian proporcionalmente en la misma dirección. Como ejemplo (no
discutido en el libro), consideremos el problema de estimar el porcentaje de
mezcla Caucásica en la población Afroamericana. Consideremos un gen. El Duffy es un buen ejemplo, y uno que ha sido utilizado
tradicionalmente. Este gen es muy frecuente en los caucásicos pero virtualmente
desconocido en los negros de África occidental. El porcentaje de este gen en la
población Afroamericana podría ser, y lo ha sido, usado para estimar el
porcentaje de mezcla caucasoide. Si un quinto de los
ancestros de la población negroide fueran caucasoides, la frecuencia del gen Duffy sería un quinto como en los caucasoides. De este
modo, a partir de un gen, el mestizaje podría ser estimado. Más generalmente
cuando dos poblaciones se mezclan, las frecuencias genéticas serían w1f1+(1-w1)f2,
donde w es el porcentaje de la
población vástago con que la primera población contribuyó, y f1 y f2
son las respectivas frecuencias genéticas. Las frecuencias genéticas están
siempre sujetas a cambios aleatorios (derivas) y a efectos de la selección
natural. Así, uno obtiene respuestas ligeramente diferentes dependiendo de los
genes estudiados. La obvia solución es examinar muchos genes y hacer un
promedio. Una vez que uno posee las frecuencias de mestizaje, uno puede
cartografiarlas en un mapa. El primer componente principal daría una buena
descripción del porcentaje de la población invasora de antaño. Este método
funcionaría incluso si uno no conociera las poblaciones que se mezclaron. Como
se mencionó, Cavalli-Sforza y asociados hicieron buen uso del mapa de un primer
componente principal para Europa, para argumentar que el patrón observado puede
ser explicado al variar las mezclas en las dos poblaciones, una original
población nómade y una población agrícola en expansión desde Oriente Próximo. Ahora supongamos
que uno tuviera tres poblaciones. Uno debiera primero computar las frecuencias
genéticas que se esperan en cada una de las poblaciones que son la mezcla en
las primeras dos. Las diferencias entre la frecuencia que podrían ser
explicadas por la mezcla de las primeras dos poblaciones, y la observada,
podrían ser atribuidas a la mezcla de una tercera. Debido a que el segundo
componente principal se construye para solo utilizar información que no está en
el primer componente principal, sus valores debieran indicar el grado de mezcla
con la tercera población. Cavalli-Sforza y
asociados aseguran haber conducido simulaciones que demuestran que los efectos
de las expansiones de antiguas poblaciones, efectivamente dejarán evidencia en
los mapas de componentes principales. Hay que notar que no habrá evidencia
escrita de una expansión de la población original, ni el nombre de aquella
población, ni sus frecuencias genéticas tendrán que ser conocidas. En numerosos casos,
los mapas de componentes principales consisten a grandes rasgos, de círculos
concéntricos, que pueden ser interpretados como indicando mestizaje con las
poblaciones circundantes, de una población original que pasó por una expansión
prehistórica. En la discusión, ellos llaman la atención sobre ciertos patrones
y se especula sobre cuáles poblaciones podrían haberse expandido. La expansión fuera
de Oriente Próximo, con la llegada de la agricultura, es un ejemplo que ha sido
discutido. Ellos interpretan un patrón de círculos concéntricos alrededor del
mar del Japón, como indicación de una expansión desde aquella área,
posiblemente de un pueblo similar a aquel de la cultura prehistórica Jomon en Japón (pág. 249). Mapas similares para Italia son
interpretados como la evidencia de la difusión de los genes de los Etruscos
originales, quienes pudieron haber tenido un patrón distintivo de frecuencias
genéticas a través de la deriva en una pequeña población original, o mediante
la inmigración a Italia desde otra área (pág. 279). El libro
proporciona mapas de componentes principales para los primeros pocos
componentes principales sobre una base mundial, y en cada capítulo continental
separado. Cavalli-Sforza y asociados han diseñado un efectivo e ingenioso
mecanismo para combinar los datos proporcionados por los principales
componentes, en mapas de colores (utilizados anteriormente en Menozzi, Piazza, & Cavalli-Sforza, 1978). El ojo humano
puede distinguir tres colores primarios, y utilizando un color primario
separado para cada primer componente principal, un mapa puede ser preparado
para que muestre los tres primeros componentes (que parecen explicar alrededor
de la mitad de la variación total en frecuencias genéticas). El resultado son
unos muy interesante mapas de colores. Uno de estos es utilizado para la
cubierta de la tapa del libro. Razas Este podría ser un
buen momento para comentar sobre las ideas de los autores sobre las razas. En
el primer capítulo hay una discusión llamada "El Fracaso Científico del
Concepto de las Razas Humanas" (pág. 19). Empieza con la declaración que
"la clasificación en razas ha resultado ser un ejercicio inútil por
razones que ya eran claras para Darwin". La referencia implícita a la idea
de de Darwin de que las razas se funden unas con otras, haciendo imposibles las
distinciones entre grupos. Los autores
recalcan que la variabilidad genética medida dentro de las poblaciones, es
mayor que la variabilidad entre las poblaciones, lo que es correcto. Sin
embargo, se equivocan al apuntar que ninguno de los genes tradicionalmente
estudiados, son los que se relacionan con tales variables como color de piel o
forma de nariz, que son las variables genéticas que muestran una mayor
variación entre las poblaciones. Es muy posible que muchos de los genes que
afectan estos rasgos se han fijado en muchas
poblaciones (juzgando por la ausencia de piel oscura entre los suecos, y la
ausencia de individuos no-albinos ligeramente pigmentados entre los
Liberianos). En este punto, al no saber qué genes influencian rasgos
socialmente significativos, no sabemos exactamente cuánto de la variación
genética en estos rasgos está entre las poblaciones, y cuánto dentro, a pesar
del que el cálculo aproximado dice que la mayoría de las variaciones
importantes están dentro de las poblaciones. El libro declara
que (pág. 19) "sin embargo, los mayores estereotipos, todos basados en el
color de la piel, el color y forma del cabello y los rasgos faciales, reflejan
diferencias superficiales que no están confirmadas por análisis más profundos
con rasgos genéticos más confiables..." Sin embargo, la evidencia en el
resto del libro sirve para desaprobar esta declaración. Ya ha sido subrayado
cómo los árboles fueron calculados, y que los diagramas de componentes
principales clasifican poblaciones de manera tal, que los grupos equivalentes a
las razas pueden ser fácilmente reconocidos. El mapa de la cubierta hace fácil
reconocer los territorios de las principales razas. Australia está en rojo,
África sub-sahariana en amarillo verdoso, Europa en verde, China, Japón, Corea
en púrpura, y el Nuevo Mundo en varias tonalidades de púrpura. Cada una de las
regiones corresponde a lo que tradicionalmente se considera como razas. Por supuesto, el
mapa efectivamente muestra intergradaciones entre los
principales grupos de poblaciones. El concepto de razas como subespecies
implica que tales graduaciones serán visibles, ya que si las poblaciones no
pudieran mezclarse serían clasificadas como especies diferentes y no meramente
razas diferentes. Otros mapas en el libro confirman la existencia de las razas.
El mapa del primer componente principal en África muestra un agudo gradiente de
norte a sur (pág. 191). Las líneas de contorno se acercan en el Sahara. Un
rápido vistazo muestra que África puede ser dividido en un área norafricana
donde viven pueblos tradicionalmente llamados caucasoides, y un África
sub-sahariana donde viven pueblos tradicionalmente llamados negroides (las dos
zonas más meridionales cogen la mayoría del África sub-sahariana). El mapa
muestra una zona en el Sahara donde las frecuencias genéticas son intermedias.
Mientras que tal zona probablemente existe en terreno, los actuales datos
genéticos para ella son débiles. Solo unos pocos grupos saharianos han sido estudiados
(los tuaregs son los más importantes). Los mapas están marcados con los puntos
de datos usados. Con mucha frecuencia, los puntos de datos son para la costa
norafricana, y para puntos al sur del Sahara. A grandes rasgos, del mismo modo
que los mapas climáticos son dibujados, el ordenador luego relleno las líneas
inconexas con zonas de suave transición. Otro mapa muy
interesante del primer componente principal, es el de Asia (pág. 250). Para
este continente, el primer componente principal explica el 35.1 % de la
variación total. Las líneas corren aproximadamente de norte a sur con valores
extremos en Oriente Próximo, y en el lejano Oriente, incluyendo Japón, China y
Vietnam. Una línea que esta entre Burma y la India
corresponde estrechamente a la tradicional división mongoloide/caucasoide en la región. Se curva para incluir al Tíbet en
el área mongoloide, y luego se dirige hacia el norte. Como el autor lo nota,
los valores más altos para el grupo caucasoide no son
adyacentes a Europa, sino a la península Arábiga, sugiriendo un posible flujo
genético desde esa área. En el lejano norte
de Eurasia (donde los datos son escasos), la línea mongoloide parece alcanzar
casi los Urales, a pesar que hay evidencia de considerable mestizaje en las
llanuras de Eurasia septentrional, en la medida que las poblaciones han migrado
en ambas direcciones. Un asunto ampliamente debatido ha sido la naturaleza de
los lapones, un grupo europeo ártico que habla un lenguaje similar al ocupado en
los Urales. Los árboles muestran el grupo lapón con otros europeos. Sus
frecuencias genéticas describen una mezcla de un 52.5 % caucásico con el resto
siendo mongoloide, a pesar que otro método muestra más mezcla europea (pág.
273). La mejor aproximación es que este grupo inmigró hacia Escandinavia desde
los Urales, trayendo un patrón mongoloide de frecuencias genéticas con ellos, y
luego se mezclaron gradualmente con otros escandinavos, hasta que sus
frecuencias genéticas obtuvieron el patrón general europeo. Los fineses, otro
grupo que habla una lengua urálica, se estima que
tienen un 90 % de genes europeos con un 10 % de urálicos,
mientras que los Húngaros (también parlantes de una lengua Urálica)
parecen tener un 10 % de mezcla no-europea (pág. 273). Hay varios otros
grupos pequeños cercanos que son difíciles de clasificar en los principales
grupos raciales. Los ainu, un pueblo tradicionalmente cazador-recolector del
norte de Japón, es notable por su apariencia caucasoide
y cuerpo velludo, prueba tener frecuencias genéticas bastante cercanas a las de
los otros japoneses, y por lo tanto debieran ser colocados dentro del grupo
mongoloide principal. Enfermedades y frecuencias genéticas La discusión
mundial termina con una sección que revisa gen por gen, con un comentario sobre
las distribuciones. Los detalles serán principalmente de valor para aquellos
interesados en un gen en particular. Esto podría ser un buen punto para
discutir si los genes estudiados son verdaderamente tan neutrales como la
teoría que yace en el libro, presume. Las distancias
medidas entre las poblaciones pueden ser reducidas si los genes en cuestión han
sido sometidos a la selección dependiente de la frecuencia. La selección
dependiente de la frecuencia ocurre cuando un gen poco común tiene una ventaja.
Un muy importante ejemplo de selección dependiente de frecuencia ocurre con los
parásitos y las enfermedades infecciosas. Las defensas del cuerpo contra
organismos foráneos dependen de la identificación de estos como foráneos, lo
que es hecho por la naturaleza de las proteínas sobre la superficie de las
células. Genéticamente, esto es controlado por el sistema antígeno humano de
los leucocitos, o sistema ALH. Hay numerosos loci,
dos de los cuales, el A y el B son muy bien estudiados en diferentes
poblaciones. Cada uno de estos loci tiene numerosos
alelos. La frecuencia de cada loci es tratada como un
"gen" diferente en este libro. De esta manera, una gran parte de la
base de datos se relaciona con estos loci. "El
más importante sistema de marcadores en nuestra colección, ALH, está
representada por 12 alelos y 17 alelos B" (pág. 130). Mientras la genética
poblacional del sistema ALH no es bien entendida, hay probablemente un grado de
selección estabilizadora. De otra forma, la variabilidad observada no habría
sobrevivido (Takahata, 1993). Los parásitos y
organismos de enfermedades evolucionan para tener proteínas que imiten aquellas
del cuerpo. El sistema inmunológico de un individuo que tiene genes ALH, que
son relativamente escasos, encontrará más fácil el reconocer los organismos
foráneos. Si cualquier alelo ALH se hace relativamente común, las enfermedades
que atacan a los portadores de ese alelo se harán más comunes (Jones, 1992,
Gráfico en pág. 287). El índice de mortalidad entre los portadores de aquel
alelo se incrementan, reduciendo la frecuencia del
alelo. Otros alelos tienen una ventaja porque su cuerpo puede reconocer mejor
los patógenos más comunes. Este mecanismo se cree que es el que promovió el
alto grado de diversidad genética que caracteriza al sistema ALH. Muchos alelos
se hallan en humanos y en especies tan diferentes como el ratón (para una
asimilable introducción del rol de los parásitos en la evolución, ver Ridley, 1994. Para una discusión más técnica sobre el
sistema ALH humano, ver Klein, 1990). Hay una breve
discusión sobre las asociaciones con la enfermedad, pero es muy posible que
haya otras asociaciones que no se conozcan, incluyendo algunas con enfermedades
que alguna vez fueron importantes pero las cuales ya no lo son. Otro sistema
genético muy importante es el ABO que es vital en la tipificación para
transfusiones de sangre. Debido a la necesidad para la tipificación sanguínea, está muy bien estudiado, y disponible para virtualmente
todas las poblaciones. Ciertos tipos sanguíneos son conocidos como más
vulnerables a ciertas enfermedades, probablemente para que el cuerpo pueda más
fácilmente reconocer ciertos organismos invasores. Por ejemplo, los individuos
O parecen relativamente resistentes a la sífilis (pág. 126). Esto podría
explicar por qué virtualmente todos los amerindios (excepto los Esquimales y ciertos grupos amerindios del norte) son tipo
O, debido a que la sífilis se cree que fue introducida en el Viejo Mundo por
Colón. Los individuos con tipo A son más vulnerables a la viruela. La
tuberculosis (pulmonar) se cree que es más virulenta en individuos A que en O ó
B. La malaria muestra preferencia por individuos A. De este modo, parece que
una selección por balance puede existir para el grupo de sangre ABO. Las frecuencias de
otros genes, se cree, son afectados por enfermedades. El alelo O Duffy (muy elevada frecuencia en africanos) confiere
resistencia a un particular parásito de la malaria, el Plasmodium
vivax. Un número de variantes de G6PD producen
resistencia a la malaria. Los genes GM y KM de la inmunoglobina,
que producen anticuerpos y juegan un importante rol en la defensa contra los
patógenos, podrían ser muy bien sujetos a una selección estabilizante. El
sistema secretor de FUT2(SE) que provoca
"secreciones de la substancia responsable por A, B y las substancias
relacionadas con H que normalmente se encuentran en las células rojas de los
individuos, y definen su estátus ABO" (pág.
133), es conocido por afectar la vulnerabilidad a las úlceras, con secretores
menos vulnerables. Como la breve
discusión anterior lo demuestra, muchos de los sistemas genéticos ampliamente
estudiados que son el sujeto de este libro, parecen posiblemente ser el sujeto
de una selección estabilizante (la selección dependiente de la frecuencia,
anteriormente mencionada), de forma que algunos escasos alelos tienen una
ventaja reproductiva. Esto tendería a reducir las diferencias genéticas entre
los pueblos del mundo. El efecto es probablemente insuficiente para hacer de la
presunción de neutralidad, que delinea gran parte de la teoría de este libro,
inaplicable. Sin embargo, el lector debe tener en mente que algunos sistemas
genéticos pueden quedar sujetos a la selección estabilizante, y otros a la
selección desestabilizante y algunos otros posiblemente a ambos. Un sistema
puede estar sujeto a ambos si las frecuencias de alelos tienden a un valor de
equilibrio particular bajo ciertas condiciones, pero esta frecuencia de
equilibrios depende de la localización. Las diferencias climáticas o culturales
podrían hacer que el equilibrio de las frecuencias genéticas dependiera de la
localización. Importantes ejemplos son los relacionados con la malaria, donde
en las áreas maláricas hay un elevado equilibrio de
frecuencias para alelos que otorgan resistencia a la malaria, y cero equilibrio
de frecuencia en regiones maláricas. Se proporciona
una útil discusión sobre la distribución de los genes que se supone protegen
contra la malaria (pág. 146-149), a pesar que esos genes no son utilizados para
calcular distancias genéticas, debido a que sus genes reflejan más selección
que deriva. Incidentalmente, el
conocimiento que muchos de los fácilmente estudiados genes aparecen como
sujetos a la selección estabilizante, es importante al evaluar un argumento comúnmente
esgrimido. Es frecuentemente aceptado que hay solo un 6.3 % variación genética
entre las razas, con el resto siendo entre poblaciones (8.3 %) o entre
individuos dentro de las poblaciones (85.4 %) (Lewontin,
1972). Aunque es probablemente cierto que la mayoría de la variabilidad
genética esté entre los individuos, las aseveraciones comunes malinterpretan la
investigación científica. Una correcta aseveración sería que un 6.3 % de la
variación de frecuencias genéticas mensurables es entre razas. El trabajo de Lewontin (citado en la pág. 19) se basa en los genes que
pudieron ser medidos en la época que él los estudio (muchos menos de los que
hoy pueden ser medidos). Ninguno de estos genes afectaba el color de piel, la
forma de nariz, la constitución física, talla, etc. para mencionar
características ffísicas que difieren entre razas.
Podemos asegurar que los genes que fueron estudiados (o pudieron ser estudiados
dado el conocimiento entonces disponible) no eran una muestra aleatoria de
todos los genes. Parece que ellos enfatizaron demasiado los genes que eran
relevantes para las defensas del cuerpo contra la enfermedad, y que estaban
sujetos a una selección estabilizante. Si esto es así, la importancia de las
diferencias raciales fue subestimada deliberadamente. Los capítulos regionales El resto del libro
está organizado del mismo modo que el capítulo Mundial, excepto que cada
capítulo enfoca sobre un área continental, y más poblaciones son discutidas
dentro de cada capítulo que las pocas de cada continente que fueron incluidas
en el estudio de las 42 poblaciones. Cada capítulo comienza con una buena
reseña de la prehistoria de la región, y una historia de las migraciones de
población hasta el 1.500 DC. Estos son útiles para los no-especialistas, pero
probablemente contienen poco que no sea conocido para los especialistas. Una matriz de
distancias es luego calculada para las poblaciones seleccionadas, y utilizada
para producir un árbol enseñando las líneas estimadas de descendencia. Esto es
discutido luego, con énfasis sobre varios cuestiones
interesantes, como el origen de poblaciones en particular. Los componentes
principales son luego calculados y discutidos. Los genes individuales también
son discutidos. Para los capítulos
sobre Asia y sobre Europa, hay un tercer nivel. Asia es discutida región por
región (Ártico, Asia nordeste, Asia sudeste, sub-continente Indio, Asia central
y Asia occidental). En Europa, a las regiones seleccionadas se les da un
tratamiento similar (Italia, Francia, península Ibérica, Cerdeña) con la
presentación de mapas de los componentes principales. Asia La mayor sorpresa
para este lector estará en el capítulo asiático. Hay una tendencia a pensar que
la tercera de las razas humanas, los chinos Han, como una población homogénea.
El análisis demuestra enormes diferencias entre el norte y el sur de China. En
un árbol con 39 poblaciones asiáticas (pág. 225), la primera separación coloca
a los chinos meridionales con otras poblaciones del sudeste asiático, como los
filipinos, malayos, thai e indonesios; siendo los thai y viet
muong los más cercanos. En contraste, el norte de
China se agrupa con Corea, Japón y el Tíbet, como podría esperarse. Sin
embargo, este grupo es efectivamente mostrado como más cercano a grupos como
los turcos, libaneses e iranís, tradicionalmente considerados como caucasoides. Una posible
explicación es que la agricultura emergió dos veces en China, una vez fue el
mijo en el norte de China y otra vez fue el arroz en el sur de China, y que
estas poblaciones luego se expandieron, congelando sus frecuencias genéticas.
La línea divisoria se ubica entre el río Yangtse y el
río Amarillo. La evidencia de respaldo es proporcionada por un análisis de una
muestra estratificada de alrededor de 540.000 apellidos chinos del Censo de 1982,
que muestra un patrón que se asegura es a grandes rasgos, similar al de las
tres áreas culturales neolíticas. La importancia de
este hallazgo de una diferencia relativamente grande entre los chinos del norte
y el sur, ya que mucha de la investigación es hecha sobre chinos nacidos en
EE.UU. y Canadá (Vernon, 1982), que son predominantemente de ancestro chino
meridional, provenientes de Hong Kong, Canton o
alrededores. Podría ser arriesgado el generalizar a partir de esto, a toda la
China Han. Para aquellos
interesados en el comportamiento y el desarrollo económico, la semejanza entre
los chinos meridionales y los filipinos, malayos, etc. presenta un problema.
Los chinos meridionales generalmente les va bien en
exámenes de inteligencia y académicos, sean hechos estos en EE.UU. o en Hong
Kong, a menudo mejor que a los caucasoides. Los filipinos generalmente no lo
hacen tan bien. En Malasia, los chinos lo hacen mejor que los malayos. Dentro
del sudeste asiático, a los chinos de ultramar generalmente les va mucho mejor
económicamente que a los malayos (Sowell, 1994). De
esta forma, es sorprendente ver las pequeñas diferencias genéticas entre los
chinos meridionales y las poblaciones adyacentes. Europa Alguien interesado
en las relaciones genéticas de variadas poblaciones encontrará mucho de interés
en los varios capítulos sobre los Continentes. Por ejemplo, al
inspeccionar el árbol para Europa (pág. 268), se descubrirá que los lapones son
la población que está más separada de otras poblaciones. Luego vienen los
sardos, que son lo suficientemente diferentes de otros europeos, como para que
su inclusión en el análisis de los componentes principales requiera que se les
dé un componente para ellos (pág. 291). Su mezcla genética única es atribuida
primariamente a la deriva genética en una población pequeña. Se descubre que
los Vascos son otro grupo distinto, quienes se argumenta son el residuo de los
europeos paleolíticos originales. Se descubre que Islandia es bastante distinta
del resto de Europa, lo que es atribuido a la deriva genética en una población
pequeña. Ninguna de estas poblaciones menores ha hecho contribuciones al
poblamiento de América. Una sección
bastante grande ubica a pueblos centroeuropeos como los ingleses, alemanes,
suecos, italianos, polacos y rusos juntos (pág. 268). Curiosamente, los
irlandeses y los escoceses están fuera de esta sección, incluso aunque muchos
piensen que ellos son muy similares a los ingleses, a lo mejor porque han
estado unidos políticamente a estos. Incluso, a pesar que hay rivalidades
históricas entre pueblos como los franceses y los alemanes, o los rusos y los
polacos, los datos aquí demuestran que cualquier diferencia genética es
demasiado pequeña para tomarla como diferencia en carácter nacional que algunos
observadores aseguraban ver. No es necesario decir que la semejanza en
frecuencias genéticas entre estos grupos de pueblos, que incluyen entre ellos a
los principales fenotipos europeas - los nórdicos, alpinos, mediterráneos - y
los eslavos, es una poderosa evidencia contra cualquier intento de superioridad
genética de los nórdicos o de los alemanes, como los nazis supuestamente
aseveraron. Es altamente improbable que los genes relevantes en el
comportamiento puedan diferir mucho en frecuencia, dadas las pequeñas diferencias
en frecuencias de los genes mensurables. La semejanza en frecuencia genética se
debe, a estas poblaciones derivaron recientemente de una o unas poblaciones
comunes (probablemente de un grupo de agricultores Neolíticos que se
diseminaron desde Oriente Próximo, seguidos de inmigrantes posteriores de las
estepas de Asia), o por un elevado nivel de mestizaje entre estas varias
poblaciones. Cualquiera de estas posibilidades sería inconsistente con las
grandes diferencias en la frecuencia de los genes más importantes socialmente,
a pesar que eso no hace tales diferencias completamente imposibles. África Una situación de
algún modo similar es hallada en África. Los antropólogos tradicionalmente
dedican mucho tiempo a pequeñas poblaciones que son interesantes, pero las
cuales constituyen relativamente pocas personas. De este modo, el capítulo
africano tiene secciones sobre los pigmeos, los khoisan y los pueblos de
Etiopía y el Sahara. Sin embargo, el grueso de la población del África
sub-sahariana está compuesto o por bantú-parlantes, o por africanos
occidentales. Las poblaciones de ambos grandes grupos, difieren poco
genéticamente unos de otros. En el caso de los bantú-parlantes, se cree que
esto se debe porque ellos surgieron de una población mucho más pequeña
originaria de Camerún. La evidencia lingüística, arqueológica e histórica para
este movimiento es obvia. La evidencia histórica es principalmente relevante
para Sudáfrica, donde la historia demuestra que los bantú
inmigraron a esta área, desplazando a los khoisan al mismo tiempo que llegaron
los europeos. La similitud en lenguajes y evidencia arqueológica traza las
primeras etapas del movimiento. Las semejanzas genéticas entre estos diferentes
grupos son consistentes con las hipotéticas migraciones, y sugiere que hubo dos
oleadas, una emigrando primero hacia el sur, y otra hacia el este, África
oriental, y luego hacia el sur (pág. 183-185). Debido a esta reciente expansión
bantú, las numerosas poblaciones bantú no difieren
mucho genéticamente unas de otras. En África
occidental, las poblaciones difieren un poco más entre ellas, pero aun así se
asemejan mutuamente. Los autores hipotetizan que esta
semejanza pudo ser provocada por una expansión desde una población única que
adoptó primero la agricultura. Una explicación alternativa provee tres
poblaciones originales, con solo la más oriental (los bantú-parlantes) en
posición para expandirse hacia el sur de África (pág. 185). De cualquier modo,
los grupos bantú y africanos occidentales no difieren mucho genéticamente. Se subrayó
anteriormente que las principales poblaciones europeas no difieren mucho unas
de otras. La mayoría de los EE.UU. está compuesta por los descendientes de las
principales poblaciones europeas, o de los descendientes de esclavos de territorio
africano occidental o bantú. Los dos grupos son bastante diferentes en
frecuencias genéticas y apariencia. En los diagramas mundiales de los componente principales, ellos están en polos opuestos
en el segundo componente principal (pág. 82). De este modo, no es sorprendente
que en América las diferencias entre los descendientes de africanos y europeos
se hayan hecho obvias, y llevaran a clasificar a la gente en dos razas, que se
ha comprobado que difieren en otros aspectos aparte de la apariencia (Herrnstein & Murray, 1994; Miller, 1994a,b,c; Rushton, 1994). Como el libro
demuestra, hay numerosas poblaciones que son intermedias en frecuencias
genéticas a aquellas otras poblaciones, como los africanos orientales, los
etíopes nilo-saharianos, los habitantes del Sahara y
los norafricanos. Hay otros grupos que de algún modo tienen patrones diferentes
de frecuencias genéticas (pigmeos, khoisan, sardos e islandeses), pero ninguno
de estos grupos contribuyó mucho al poblamiento de EE.UU. Se puede argumentar
que hay clinés (poblaciones intermedias) en el Viejo
Mundo, con frecuencias genéticas que cambian gradualmente de norte a sur (a
pesar que lo hacen relativamente rápido a través del Sahara). Esto no altera el
hecho que la vasta mayoría de los ancestros (no-mongoloides) de la población de
EE.UU. puedan ser clasificados como negroides (bantú o africano occidental) o caucasoide (europeo). Por supuesto, han ocurrido mezclas
posteriores, y hay muchos norteamericanos cuyo ancestro es hoy mixto. Aparte de las
principales poblaciones de África, hay numerosos grupos de poblaciones menores
que son de interés. El libro está lleno con fascinantes hallazgos sobre estas
poblaciones. Los tuareg, quienes han sido siempre un pueblo muy móvil (pág.
173) se extienden sobre un área que abarca los confines septentrionales del
árido Sahara (Argelia y Libia) hasta el Sahel (pág.
171). Los autores (pág. 173) demuestran que hay un sorprendente grado de
semejanza genética entre los tuareg y los beja (cuya
distancia genética de los tuareg es solo 135), un pueblo del Sahara oriental,
cuyo territorio limita con el mar Rojo. Debido a que cada
lector debe encontrar al menos un error, podría hacerse notar que la
localización de los Beja es diferente en el mapa de la
página 170 al de la página 171 (el cual es probablemente el correcto). La similitud
genética es sorprendente dada la gran distancia geográfica. Los autores hipotetizan sobre un origen común, posiblemente unos 5 mil
años atrás. Esto es un tiempo largo, pero el mínimo que la migración este-oeste
a través del Sahara requirió para los grupos que tuvieron un origen común, es
mucho mayor que el ancho del Sahara. Unas pocas de esas migraciones hace
alrededor de decenas de miles de años pudieron reducir de gran manera o
eliminar cualquier diferencia genética entre Noráfrica
y África sub-sahariana. Pero, como los autores lo documentan muy bien, la
diferencia genética entre los habitantes caucasoides de Noráfrica
y los habitantes negroides del sur, en África occidental, es bastante grande
(para no mencionar las obvias diferencias en color de piel y otros aspectos de
la apariencia). Esto hace muy posible que las actuales poblaciones norafricanas
no evolucionaran en el lugar sino que llegaran desde otro sitio, debido a que
si lo hubieran hecho no serían tan diferentes de los otros africanos como lo
son. De esta manera, las
grandes diferencias genéticas al norte y sur del Sahara presentan un problema
que no es fácilmente resuelto meramente notando que hay una baja densidad, con
un área seca en medio, debido a que grandes migraciones de población (trayendo
genes con ellas) aparentemente han ocurrido. He desarrollado una
teoría (Miller, 1994d) en que las grandes diferencias genéticas entre las
poblaciones euroasiáticas y las africanas, que Cavalli-Sforza y asociados tan
bien han documentado, se deben parcialmente a un movimiento temprano moderno
fuera de África, hacia Eurasia, seguido por el movimiento de los Neanderthal
hacia Oriente Proximo. Esto dividió a la población
moderna en dos grupos. Más tarde, una rama, o ramas de la población caucasoide europea inmigraron a Noráfrica. Hay una enorme área
de África cuya afinidad genética ha sido poco clara. Me refiero a Etiopía y
áreas adyacentes. La gente de esas regiones tiende a tener rasgos en cierto
modo caucasoides pero pieles oscuras. Los datos de frecuencias genéticas
sugieren que los amhara (el grupo etíope dominante) tienen frecuencias
genéticas creadas por una mezcla de 57 % de genes africanos nilóticos con 43 %
de genes de norafricanos (pág. 174). Otras poblaciones europeas son similares.
De este modo, si uno debe clasificar a estos pueblos en una raza principal,
ellos debieran ser denominados negroides. La historia registrada de la región y
su localización hace muy posible que hubiera ocurrido una efectiva mezcla de
pueblos caucasoides y negroides allí. Otro grupo que ha
sido sujeto de mucha discusión son los pueblos khoisanidos
(incluyendo a los hotentotes, san, 'kung). Los san
(bosquimanos) en el sur de África son un grupo que físicamente luce bastante
diferente a otros negroides. Baker (1974) y Coon
(1965), entre otros, han aseverado que ellos son tan diferentes de los
negroides, como los caucasoides lo son, y debieran ser tratados como una raza
separada de otros negroides. Los datos genéticos reportados aquí demuestran que
ellos difieren más de otros africanos sub-saharianos, que lo que otros grupos
sub-saharianos difieren entre sí (pág. 175). Es de interés notar
que los san son más cercanos a las poblaciones de Oriente Proximo,
que a las poblaciones bantú adyacentes. Sus frecuencias genéticas son
consistentes con un casi 56 % de Oriente Proximo, y
el resto es africano. Dado que el territorio que ellos actualmente ocupan está
muy distante del territorio caucasoide, esto es
asombroso. Sin embargo, una posible teoría, respaldada por restos históricos y
trazos lingüísticos, sostiene que ellos alguna vez estuvieron en África
oriental, posiblemente tan al norte como Egipto. Debieron haber tenido una
cierta mezcla con caucasoides. Las poblaciones
etíopes, que parecen ser una mezcla caucasoides/negroide similar, muestran una
considerable distancia genética de los san, sugiriendo que si ambos son el
resultado de una mezcla, estas ocurrieron en épocas diferentes. Una hipótesis
alternativa, que es respaldada por la evidencia del ADN mitocondrial y la
distintiva morfología de los san, es que ellos son una
población residual de una antigua raza de humanos cuyos territorios alguna vez
cubrieron gran parte de África, y que son los ancestros de todos los humanos
(pág. 176). Es interesante ver como los modernos datos genéticos respaldan la
temprana idea de Coon, de que aquellos eran una
reliquia de las poblaciones originales, desde la cual otros grupos se separaron
(1963-65). Así, la semejanza con las poblaciones de Oriente Proximo
es explicada debido a que estas se derivan de los san. Desafortunadamente,
hay muy pocos datos con frecuencias genéticas para Madagascar, y esta isla es
frecuentemente dejada fuera de los mapas debido a la falta de información.
Madagascar es potencialmente muy interesante debido a que el lenguaje de los
malgaches es similar a los lenguajes de Borneo sud-central. Generalmente se
cree que Madagascar fue colonizado por un pueblo de origen austronesio,
más que de la cercana África (pág. 168), cuyos habitantes aparentemente no
habían desarrollado naves apropiadas. América "La evidencia
genética para América confirma absolutamente la migración de tres oleadas
sugerida por la evidencia dental y lingüística: amerindios, na-dene y eskimos" (pág. 349).
Por supuesto, mayores e interesantes detalles son proporcionados por el
capítulo. Por ejemplo, el elevado grado de diversidad genética entre las tribus
sudamericanas se atribuye a la deriva en numerosas poblaciones pequeñas. Australia, Nueva
Guinea y las islas del Pacífico son discutidas en el capítulo final. La
evidencia genética no es particularmente definitiva para Australia y Nueva
Guinea. El libro concluye
con un llamado para una mayor investigación, y para la recopilación de datos
sobre numerosas poblaciones pequeñas del mundo, antes que desaparezcan. Tal
esfuerzo está encaminado como parte del proyecto del Genoma Humano. Conclusiones Para el estudiante
de las poblaciones humanas este libro aclara numerosos puntos. Uno es que hay
una considerable variabilidad genética entre las poblaciones. Los grupos
humanos difieren en mucho más que en cantidad de melanina. Esto hace más
plausible que ellos difieran socialmente, de modo importante, incluyendo
inteligencia, personalidad, resistencia a las enfermedades, comportamiento
sexual, etc. Mientras otros
pueden mencionar la existencia grupos pequeños, parece haber tres grupos
principales humanos que incluyen a la mayoría de las poblaciones humanas, y
cuyas frecuencias genéticas difieren. Esos son los tres grupos tradicionales
llamados negroides, caucasoides y mongoloides. Los amerindios y australianos
constituyen otros grandes grupos con frecuencias genéticas distintivas. En fin, este es un
libro muy valioso que debiera estar en toda biblioteca universitaria, a pesar
que su elevado precio lo tendrá fuera de la mayoría de las librerías privadas. Referencias 1.
Ammerman, A. J. & Cavalli-Sforza, L. L. (1973). A
population model for the diffusion of early farming in Europe. In The
Explanation of Culture Change. C. Renfrew ed. pp. 343-357. London: Duckworth.
2.
Austen, R. A. (1979). The trans-Saharan slave trade: A
tentative census. In The Uncommon Market: Essays in the Economic History of the
Atlantic Slave Trade. Gery, H. A. & Hogendorn,
J. S. New York: Academic Press
3.
Baker, J. R. (1974). Race. Oxford: Oxford University
Press. 4.
Barbujani, G., Pilastro,
A., Domenico, S. D., & Renfrew (1994). Genetic
variation in North África and Eurasia: Neolithic demic diffusion vs.
Paleolithic colonization. American
Journal of Physical Anthropology 95,
137-154. 5.
Bouchard, T. J., Lykken, D. T., McGue, M., Segal, N.
L., & Tellegen, A. (1990). Sources of human psychological differences: the
Minnesota study of twins reared apart. Science, 250, 223-228. 6.
Brues, Alice M. (1990). People and Races. (Prospect
Heights, Illinois: Waveland Press). 7.
Cavalli-Sforza,
L. L., Menozzi, P., & Piazza, A. (1994). The
History and Geography of Human Genes. Princeton:
Princeton University Press.
8.
Clark, J. D. (1993). Áfrican and Asian perspectives on
modern human origins. in Aitken, J., Stringer, C. B., & Mellars, P. A. Eds.
The Origin of Modern Humans and the Impact of Chronometric Dating. Princeton: Princeton University
Press. pp. 148-178. 9.
Coon, C. S. (1963). The Origin of Races. New York:
Alfred A. Knopf. 10. Coon, C.
S. (1965). The Living Races of Man. New York: Alfred A. Knopf. 11. Coon, C.
S. (1982). Racial Adaptations. Chicago: Nelson-Hall. 12. Eaves, L.
J., H. J. Eysenck, & N. G. Martin.1989. Genes, Culture, and Personality.
London: Academic Press. 13. Edwards,
A. W. F. & Cavalli-Sforzaa, L. L. (1964). Reconstruction of evolutionary
trees. In Phenetic and Phylogenetic Classification. V. E. Heywood and J.
McNeill, eds. pp. 67-76. London: The Systematic Association. 14.
Hammer, Michael F. (1994). A recent insertion of an
Alu element on the Y chromosome is a useful market for human population
studies. Molecular Biology and Evolution,
11, 749-762. 15.
Harpending, H. C., (1994). Signature of ancient
population growth in a low-resolution mitochondrial DNA mismatch distribution. Human Biology, 66,
591-600. 16.
Harpending, H. C., Sherry, S. T., Rogers, A. R. &
Stoneking, M. (1993). Genetic structure of ancient human populations. Current Anthropology 34, 483-496. 17.
Herrnstein, Richard J. & Murray, Charles (1994).
The Bell Curve: Intelligence and Class Structure in American Life. New York: The Free
Press. 18. Jones, S.
(1992). Natural selection in humans. In The Cambridge Encyclopedia of Human
Evolution. Jones, S., Martin, R., & Pilbeam, Eds. Cambridge: Cambridge
University Press. 19.
Kingdon, Jonathan (1993). Self-Made Man. London: Simon
& Schuster. 20. Klein,
Jan (1990). Immuniology. Boston: Blackwell Scientific. 21. Krantz,
G. S. (1980). Climatic Races and Descent Groups. North Quincy: Christopher
Publishing House. 22. Laska-Mierzejewska,
T. (1982). Dymorfizm Plclowy Czlowieka Odmiany Bialej I Czarnej Na Kubie,
Warsaw. 23.
Lee, R. B. (1968). What hunters do for a living. In
Lee, R. B. & DeVore, I. Man the Hunter. Chicago: Aldine, 30-48. 24.
Lewontin, R. C. (1972). The apportionment of human
diversity. In Evolutionary Biology, Vol. 6, T. H. Dobzhansky, M. K. Hecht, and
W. C. Steere, eds. pp. 381-398. New
York: Appleton-Century-Crofts. 25.
Loehlin, J. M. (1992). Genes and Environment in
Personality Development. Newbury
Park: Sage. 26. Lynn, R.,
(1991a). Race differences in intelligence: a global perspective, Mankind
Quarterly, 32, 254-296. 27. Lynn, R.,
(1991b). The evolution of racial differences in intelligence, Mankind
Quarterly, 31, 99-121. 28.
Menozzi, P., Piazza, A., & Cavalli-Sforza, L,
(1978). Synthetic maps of human gene frequencies in europeans? Science, 201, 786-792. 29. Miller,
E. M, (1993). Could r selection account for the Áfrican personality and life
Cycle, Personality and Individual Differences 15, 665-676. 30. Miller,
E. M. (1994a). Paternal provisioning versus mate seeking in human populations,
Personality and Individual Differences, 17, 227-255. 31.
Miller, E. M. (1994b). Optimal Adjustment of Mating
Effort to Environmental Conditions: A Critique of Chisholm's Application of
Life History Theory, with Comments on Race Differences in Male Paternal
Investment Strategies. Mankind Quarterly, XXXIV
(Summer) No. 4. 32. Miller,
E. M. (1994c).Some Implications of Bayes Theorem for Personnel Selection,
Journal of Social, Political, and Economic Studies (in press). 33. Miller,
E. M. (1994d). Did the Neandertals Separate the Áfricans from the Eurasians?
unpublished manuscript. 34.
Nei, M. & Livshits, G. (1989). Genetic
relationships of european, Asians, and Áfricans and the origin of modern Homo
sapiens. Human Heriditary, 39, 276-281. 35. Nei, M.
& Roychoudhury, A. K. (1993). Evolutionary relationships of human populations
on a global scale. Molecular Biology and Evolution, 10, 927-943. 36.
Pearson, R. (1991). Race, Intelligence and Bias in
Academe. Washington:
Scott-Townsend. 37.
Relethford, J. H. & Harpending, H. C. (1994).
Craniometric variation, genetic theory, and modern human origins. American Journal of Physical Anthropology,
95, 249-270. 38.
Ridley, Matt (1994). The Red Queen. New York: MacMillan.
39.
Rowe, D. C. (1994). The Limits of Family Influence:
Genes Experience and Behavior. (New York: Guilford: 1994). 40.
Rushton, J. P., (1994b). Race, Evolution and Behavior:
A Life History Perspective. New
Brunswick: Transaction Publishers.
41.
Sowell, T. (1994). Race and Culture: A World View. New York: Basic Books.
42.
Takahata, N. (1993). Evolutionary genetics of human
paleo-populations. In Mechanisms of Molecular Evolution. Takahata, N.and Clark,
A. G. (ed.) Sunderland, Mass.: Sinauer Associates. 43.
Torroni, A., Semino, O., Scozzari, R., Sirugo, G.,
Spedini, G., Abbas, N., Fellous, M., & santachiara Benerecetti, A.S.
(1990). Y chromosome DNA polymorphisms in human populations: differences
between Caucasoids and Áfricans detected by 49a and 49f probes. Annals of Human Genetics, 54, 287-296. 44.
Vernon, P. E. (1982). The Abilities and Achievements
of Orientals in North America. New
York: Academic Press. Notas a píe de página
·
Miller, Edward M, "Environmental Variability
Selects for Large Families only in Special Circumstances: Another Objection to
Differential K Theory," Personality and Individual Differences, Vol. 19
(December 1995), No. 6, 903-918. ·
Miller, Edward M, "Race, Socioeconomic Variables,
and Intelligence: A Review and Extension of The Bell Curve," Mankind
Quarterly, Vol. XXXV, (Spring
1995), No. 3, 267-291. ·
Miller, Edward M. and Martin, Nicholas G.
"Analysis of the Effects of Hormones on Opposite-Sex Twin
Attitudes?," Acta Geneticae Medicae et Gemellologiae: Twin Research, Vol.
44, No. 1, 1995, 41-52.
·
Miller, Edward M, "Reported Myopia in Opposite
Sex Twins: A Hormonal Hypothesis," Optometry and Vision Sciences , Vol. 72,
(January 1995) No. 1, 34-36. ·
Miller, Edward M, "Intelligence and Brain
Myelination: A Hypothesis," Personality and Individual Differences, Vol
17, (December 1994) No. 6, 803-833. ·
Miller, Edward M, "Tracing the Genetic History of
Modern Man," Mankind Quarterly, Vol. 35 (Winter 1994) No. 1-2, 71-108. ·
Miller, Edward M, "The Relevance of Group
Membership for Personnel Selection: A Demonstration Using Bayes Theorem,"
Journal of Social, Political, and Economic Studies Vol. 19 (Fall 1994) No. 3,
323-359. ·
Miller, Edward M, "Prenatal Sex Hormone Transfer:
A Reason to Study Opposite-sex Twins," Personality and Individual
Differences, Vol. 17, October 1994, No. 4, 511-529. ·
Miller, Edward M, "Paternal Provisioning versus
Mate Seeking in Human Populations," Personality and Individual
Differences, Vol. 17, August 1994, No. 2, 227-255. ·
Miller, Edward M, "Optimal Adjustment of Mating
Effort to Environmental Conditions: A Critique of Chisholm's Application of
Life History Theory, with Comments on Race Differences in Male Paternal
Investment Strategies." Mankind Quarterly, XXXIV (Summer 1994)
No. 4, 297-316. ·
Miller, Edward M, "The Consistency of Leontief
Production Functions with Perfect Substitutability Between Factors,"
Journal of Financial Management and Analysis., Vol. 7, January-June, 1994, No.
1, 35-43. ·
Miller, Edward M, "Liquidity: Its Origins and
Implications in an Uncertain Multiperiod World with Limited Borrowing." The American Economist, Vol. XXXVIII, Spring 1994, No. 1, 36-46. ·
Miller, Edward M, "Could r Selection Account for
the Áfrican Personality and Life Cycle." Personality and Individual Differences, Vol. 15, December
1993, No. 6, 665-676. ·
Miller, Edward M, "Equivocation in Mathematical
Economics Arguments," The American Economist, Vol. XXXVII, Fall 1993,
No. 2, 62-66. ·
Miller, Edward M, "An Analysis of Quality
Adjusted Price Indices and Growth Accounting: An Appraisal of the Solow Vintage
Model," Journal of Financial Management and Analysis., Vol. 6,
July-December 1993, No. 2, 58-71. ·
Miller, Edward M, "Firm Size Related Implications
of the Cost of Accounting Information and Analysis," Review of Financial
Economics, Vol. 1, Spring 1992, No. 2, Spring 1992, 68-80. ·
Miller, Edward M, "On the Correlation of Myopia
and Intelligence," Genetic, Social, and General Psychology Monographs,
Vol. 118, No. 4, November 1992, 363-383. ·
Miller, Edward M, "Ricardian Rent, Factor Quality
Variations, and the Testable Implications of Production Function
Regularity." The Review of Political Economy,
Vol. 4, No. 4, October 1992, 467-483. ·
Miller, Edward M, "Is Aggregation of Capital by
its Rent Reasonable? Implications for Growth Accounting," Journal of
Financial Management and Analysis, Vol. 5, No. 1, January-June 1992, 33-38.![]()
{kind=link}
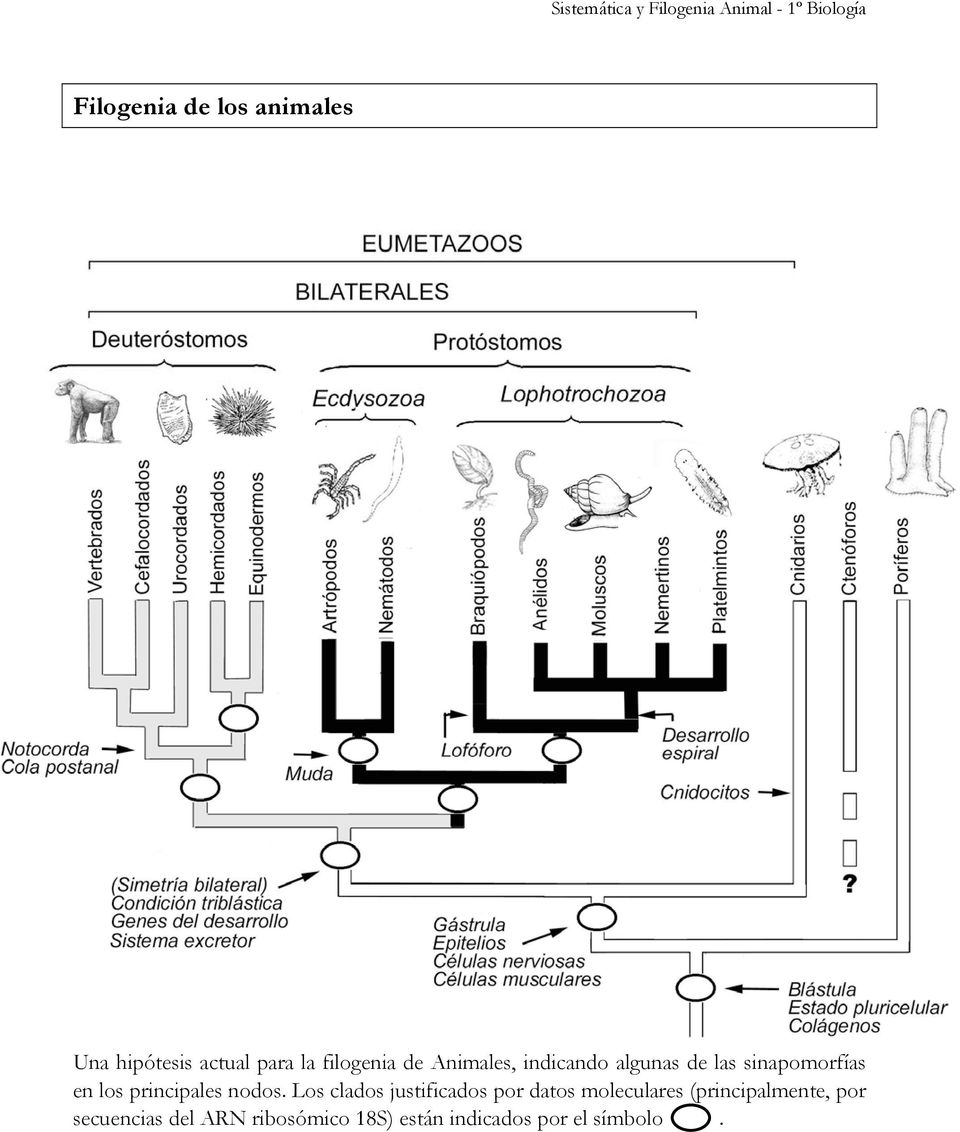{kind=link}
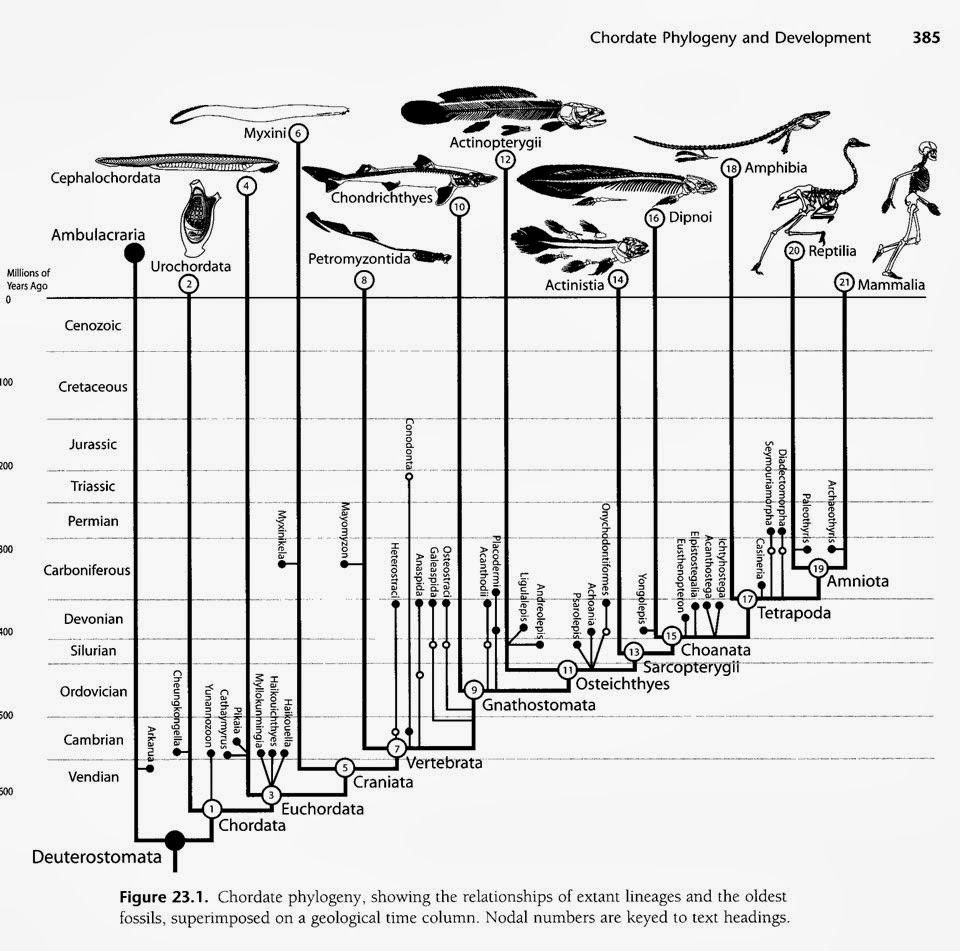{kind=link}
{kind=link}
{kind=link}
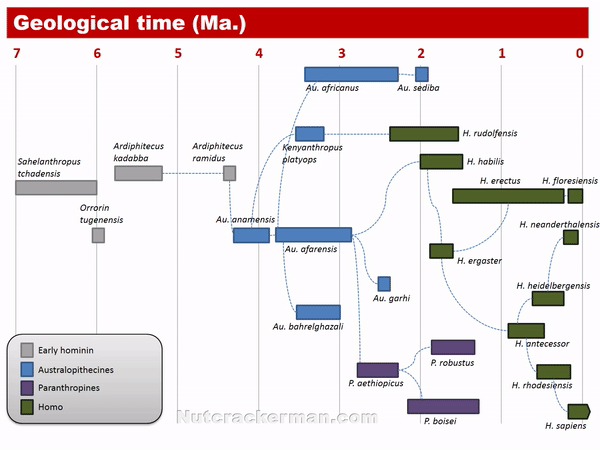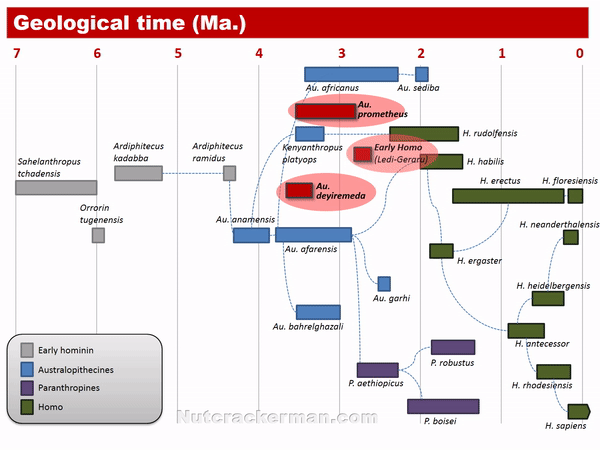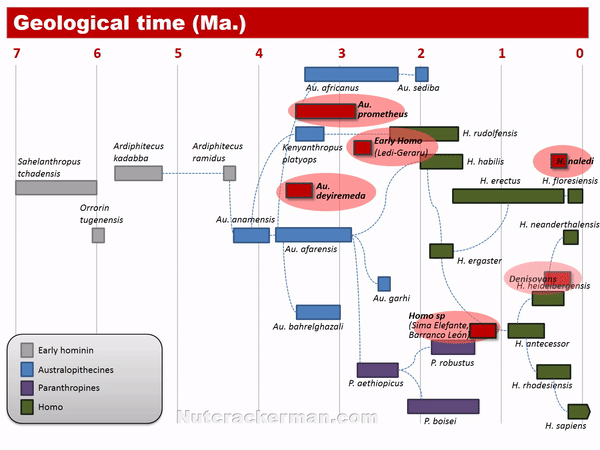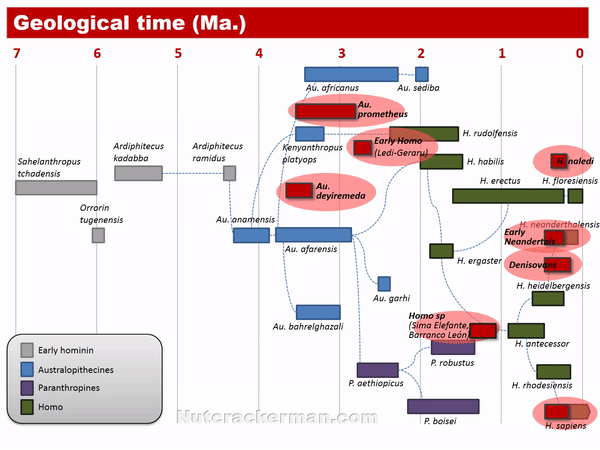{kind=link}
{kind=link}
{kind=link}